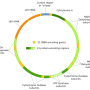
Credit: Wikipedia
Although the CRISPR/Cas9 system has seen widespread application in editing the nuclear genome, using it to edit the mitochondrial genome has been problematic. The main hurdles have been a lack of suitable editing sites in the small mtDNA, and the traditional difficulty of importing the guide RNA into the mitochondrial matrix where nucleoids can be accessed.
Two recently published papers suggest that significant progress is being made on both fronts. The first paper, published in the journal SCIENCE CHINA Life Sciences, used CRISPR techniques to induce insertion/deletion (InDel) events at several mtDNA microhomologous regions. These InDel events were triggered specifically by double-strand break (DSB) lesions. The authors found that InDel mutagenesis was significantly improved by sgRNA multiplexing and a DSB repair inhibitor called iniparib, suggesting a rewiring DSB repair mechanisms to manipulate mtDNA. In the second paper, published in the journal Trends in Molecular Medicine, the researchers give a global overview of recent advances in different forms of nuclear and mitochondrial genome editing.
To gain more insight into some of these new developments, I reached out to Payam Gammage, an expert in mitochondrial editing with a proven track record in perfecting a slightly different editing technology based on zinc finger nucleases (ZFNs). These nucleases are able to target double-stranded mitochondria for cleavage at precise base pair locations, and can therefore eliminate heteroplasmic mitochondria that have faulty nucleoids. More recently, Payam has discovered that 25 of the 30 most mutated genes found in cancer are found in mtDNA. These mutations occur at specific loci in about 60% of all tumors and, at least in colorectal cancer, actually prolong patient lifespan by ~nine years compared to wtDNA. Over 70% of colorectal cancers have at least one mtDNA which is found at heteroplasmy levels higher than 5%.
While nucleases can edit out deleterious mutations by selecting for the right mitochondria, a technology that can edit-in new variants, so to speak, is something yet to be perfected. While the methods for CRISPR editing described in the above papers sound promising, Payam related three major concerns that make take some of the wind out of their sails.
The first point is that the Life Sciences paper does not fully address the issue of targeting sgRNA to mitochondria. Secondly, a very low level double-strand break religation has previously been described in mammalian mitos. Cas9 protein expressed at high levels without gRNA results in nonspecific double-strand induction. And thirdly, the DSB repair inhibitor the researchers used may not actually do what has been traditionally thought. In other words, although it was once believed to inhibit PARP (Poly (ADP-ribose) polymerase), it was later shown to operate on different pathways. Furthermore, PARP is not even localized to mitochondria.
An interesting new approach to precise, nondestructive mitochondrial editing that does not require CRISPR techniques was recently discovered by David Liu from Harvard and MIT's Broad Institute. You may not recognize his name even though he has often been cited as the actual inventor of CRISPR, because the higher powers over at the modern and progressive Nobel Committee deemed he did not fit the bill. Liu's method relies on a bacterial toxin, DddA, that catalyzes deamination of cytosines within double-stranded DNA. By adding in a uracil glycosylase inhibitor and TALEN-like proteins, Liu created RNA-free DddA-derived cytosine base editors (DdCBEs) that can catalyze C•G-to-T•A conversions in human mtDNA with high target specificity and product purity.
To further explore the potential of DdCBEs, Liu's group successfully edited five mitochondrial genes: MT-ND1, MT-ND2, MT-ND4, MT-ND5 and MT-ATP8. Anyone who would like to get a hand on some of this technology can access the plasmids that Liu has uploaded to Addgene. For example, there is an ND4 construct on the site called ND4-DdCBE-right side TALE, which has a pCMV backbone and is expressed in mammalian cells. While complete mitochondrial editing of specific base pairs is far superior to simply cleaving mtDNA, the full generality of the approach remains to be seen. Correction of mutants will only be feasible if the faults lie within the specific conversions the editor can perform.
Having this kind of technology on tap does raise the question of whether or not new and beneficial forms of persistent mitochondrial heteroplasmy can be created. For example, it may be possible to introduce or create somatically heteroplasmic mitochondria that are better adapted to high-altitude oxygen levels, or that have enhanced thermogenesis. In any case, it would be unlikely that these manipulations could ever be inherited; if not, they cannot find their way into the germ cells. Three-parent embryo champions aside, artificially introducing or otherwise modifying the mitochondria inside the egg would be the last (and most dangerous) place anyone should begin clinically mucking around.
A curious vertebrate known as the Tuatara was recently found to maintain two independent lineages of mitochondria despite sequence divergence of around 10%. This is quite unheard-of in the animal kingdom, save for a few bivalve mollusks that are known to have biparental inheritance of distinct male and female mitochondria. Major differences were reported for control regions and origins of replication in the the Tuataran mtDNA. Researchers suggest that having two divergent mt genomes may confer an adaptive advantage for an unusually cold-tolerant reptile.
In humans, there is abundant need for mitochondrial editing for several neurologic and rare diseases. For example, autism has been associated with a G8363A transfer RNA(Lys) mutation. Other studies have recently demonstrated a mitochondrial deficiency involving an ND6 gene missense mutation (ND6P25L) that results in mice with decidedly autistic endophenotypes. ND6 is a subunit of NADH dehydrogenase that forms part of respiratory complex I. Although autism is notoriously fraught with inconsistencies in trying to nail down causative genes from nuclear GWAS studies, mitochondrial editing in animal models may be a more direct way to better define, and ultimately cure, many of these ailments that have a significant underlying mitochondrial component.
More information: Bang Wang et al. CRISPR/Cas9-mediated mutagenesis at microhomologous regions of human mitochondrial genome, Science China Life Sciences (2021). DOI: 10.1007/s11427-020-1819-8
Jiameng Dan et al. Expanding the Toolbox and Targets for Gene Editing, Trends in Molecular Medicine (2021). DOI: 10.1016/j.molmed.2020.12.005
Journal information: Science China Life Sciences
© 2021 Science X Network