New study hints that stem cells prepare for maturity much earlier than anticipated
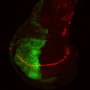
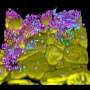
 and Pol II (12 nm gold particles) antibodies. Credit: Courtesy of Shilatifard Lab, Stowers Institute for Medical Research.")
Unlike less versatile muscle or nerve cells, embryonic stem cells are by definition equipped to assume any cellular role. Scientists call this flexibility "pluripotency," meaning that as an organism develops, stem cells must be ready at a moment's notice to activate highly diverse gene expression programs used to turn them into blood, brain, or kidney cells.
Scientists from the lab of Stowers Investigator Ali Shilatifard, Ph.D., report in the December 27, 2012 online issue of Cell that one way cells stay so plastic is by stationing a protein called Ell3 at stretches of DNA known as "enhancers" required to activate a neighboring gene. Their findings suggest that Ell3 parked at the enhancer of a developmentally regulated gene, even one that is silent, primes it for future expression. This finding is significant as many of these same genes are abnormally switched on in cancer.
"We now know that some enhancer misregulation is involved in the pathogenesis of solid and hematological malignances," says Shilatifard. "But a problem in the field has been how to identify inactive or poised enhancer elements. Our discovery that Ell3 interacts with enhancers in ES cells gives us a hand-hold to identify and to study them."
In 2000, Shilatifard identified Ell3 as the third member of the Ell (for "Eleven-nineteen lysine-rich leukemia gene") family of elongation factors, proteins that increase the rate at which genes are expressed. "At the time, we didn't think much of Ell3 because it was highly expressed in testes," says Shilatifard, noting that then people thought that sperm were merely vessels used to carry paternal DNA to an egg and that associated factors would have little relevance to the regulation of future gene expression in the resulting embryo.
But a few years back, a curious Open University graduate student working in the Shilatifard lab, Chengqi Lin, started exploring a potential function for the neglected gene by initiating a global search for regions occupied by Ell3 in the genome of mouse embryonic stem cells. His search in collaboration with a bioinformatician in the Shilatifard lab, Alexander S. Garruss, revealed that Ell3 sits on more than 5,000 enhancers, including many that regulate genes governing stem cell maturation into spinal cord, kidney, and blood cells.
"What was interesting was that Ell3 marked enhancers that are active and inactive, as well as enhancers that are known as "poised," says Lin, referring to a transition state from inactive to active. "That indicated that Ell3's major function might be to prime activation of genes that are just about to be expressed during development."
The fact that silent genes can be "primed" for expression was no surprise: researchers knew that the enzymatic machine that copies DNA into the RNA blueprint for proteins—a protein called Pol II—often pauses at the start of a gene, presumably revving its engine in preparation to jump across the genetic start gate in response to a developmental signal. However, Shilatifard and colleagues showed several years ago that paused Pol II is not a prerequisite for rapid transcriptional induction.
The surprise came when researchers used a molecular trick to deplete mouse ES cells of Ell3 and then did a "genomic" survey. They found that paused Pol II vanished from the start sites of many genes in Ell3-deficient cells. This means that not only does Ell3 preferentially mark stem enhancers, but also that its presence there is necessary to keep an idling Pol II ready for action.
Most of the current study defines how, when the developmental time is right, enhancer-bound Ell3 cooperates with components of a big-boss elongation factor called the Super Elongation Complex to release Pol II from the start gate, allowing the expression of genes required for stem cell differentiation. Critical among those findings is their observation that mouse stem cells depleted of Ell3 failed to activate genes expressed in mature cell types.
These results alone are cause for any lab to start chilling the champagne, yet a surprising coda to the study, leaves readers with yet another revelation. Collaborating with Fengli Guo, Ph.D., head of the Stowers electron microscopy core, the team prepared highly magnified images of mouse sperm and observed that both Ell3 and Pol II were present, in sperm nuclei.
In mammals, gene expression regulated by Pol II, a process known as transcription, does not begin until the formation of a single-celled zygote, that is, well after the union of sperm and egg germ cells. "It is very significant that Ell3 and other factors that regulate transcription are found in sperm," says Lin, the study's first author. Lin is cautious in interpreting this finding, "but it would be very exciting to further investigate whether transcription factors found in sperm could contribute to the decondensation of sperm chromatin or even further gene activation after fertilization by serving as epigenetic markers."
Shilatifard is also cautious as questions remain to be explored, among them whether Ell3 and Pol II actually contact DNA inside sperm or whether similar processes occur in unfertilized eggs and function in this process. Nonetheless, he feels this finding has fundamental implications, not only for development, but also for where he's going next.
"This work has opened up a whole new area of research in my lab," says Shilatifard, who has in the last decade focused on aberrant gene expression associated with leukemia. "If we find that transcription factors bind to specific regions of chromatin in germ cells, I may focus on germ cells in the next few decades. This would open a huge door enabling us to determine the role of these factors in early development."
Journal information: Cell
Provided by Stowers Institute for Medical Research