Transcriptional elongation control takes on new dimensions
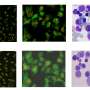
. Ice1, a subunit of the LEC (green) is associated with snRNA genes on Drosophila polytene chromosomes. Ser2 phosphorylated Pol II is shown in red. Courtesy of the Shilatifard lab, Stowers Institute for Medical Research")
Life is complicated enough, so you can forgive the pioneers of DNA biology for glossing over transcriptional elongation control by RNA polymerase II, the quick and seemingly bulletproof penultimate step in the process that copies the information encoded in our DNA into protein-making instructions carried by messenger RNA.
In a new report appearing in the Dec. 23, 2011, issue of Molecular Cell, researchers at the Stowers Institute for Medical Research add not just a new layer, but a whole new dimension to transcriptional elongation control with evidence that for each class of genes transcribed by RNA polymerase II (Pol II), there exists a specific class of elongation factors.
The Stowers team, led by investigator Ali Shilatifard, Ph.D., discovered that ELL, short for eleven-nineteen lysine-rich leukemia, not only belongs to an assemblage of transcription elongation factors, which Shilatifard's lab had identified as the "Super Elongation Complex" (SEC) a few years ago, but also that ELL is part of a distinct "Little Elongation Complex" (LEC), which acts on a completely different class of genes transcribed by Pol II. Their findings illustrate that the elongation stage of transcription is a much more specific regulator of gene expression than previously believed.
"About fifteen years ago, transcriptional elongation control was not considered all that important for the regulation of gene expression," says Shilatifard of the standard biology textbook descriptions of RNA transcription, which assume that the molecular machinery that supported transcription elongation was one-size-fits-all. "Once RNA polymerase II departed from the promoter regions, it didn't matter all that much what happened next," he says.
Transcriptional elongation is the step following promoter clearance and the step before termination, and was considered to be largely unregulated. The old metaphor was a train running on tracks. "Polymerase is the train. It sat at the promoter—which would correspond to the station," Shilatifard explains. "The polymerase train would leave the promoter/station and before long would arrive at the end of the gene. The process of the train traveling between the station and the endpoint of the gene—was considered elongation."
The latest findings derail the train metaphor. "We have shown that there are specific classes of elongation factors for different classes of genes. Therefore, much more is involved than a train simply following a predestined track," he says. "Years ago, B.F. Goodrich (the tire company) advertised that, 'for every road, there is a tire'. What we are learning is that for every class of genes, there seems to be a specific class of elongation factors. The specificity of the complexes seems to control which classes of genes are transcriptionally regulated", says Shilatifard.
Edwin Smith, Ph.D., a research scientist in Shilatifard's lab, identified LEC in Drosophila cells while biochemically dissecting the proteins associated with the Drosophila homolog of the ELL protein. In human cells, where ELL is found within the SEC, it is required to induce the expression of a class of genes specific for the pathogenesis of a subtype of genes involved in acute leukemia.
This type of leukemia results when, through a process known as translocation, the mixed lineage leukemia (MLL) gene becomes fused to any of a number of seemingly unrelated genes. In earlier studies, Shilatifard's group found that many of MLL's fusion partners, including ELL, belong to the SEC. When MLL fuses with any of these unrelated partners, the whole SEC, much like an entourage, now follows MLL to its normal target genes misregulating their elongation and ultimately causing leukemia.
While humans have three ELL genes, fruit flies have only one ELL, but its structural similarity to the human ELLs suggested an evolutionarily conserved and vital function. To find out more about ELL's function in both creatures, Smith searched for Drosophila interaction partners in collaboration with Michael Washburn, Ph.D., and Laurence Florens, Ph.D., who head proteomics at the Stowers. They used their multi-dimensional protein identification technology, or MudPIT, to identify a set of relatively uncharacterized proteins in Drosophila that associate with the C-terminus of ELL in a complex the Shilatifard lab named the "Little Elongation Complex" or LEC.
When Smith knocked down LEC subunits in fruit flies and analyzed the global expression pattern defect with Alexander Garrett, Ph.D., a bioinformatician in the Shilatifard lab, they learned that the expression levels of small nuclear RNA (snRNA) genes plummeted. Unlike other RNAs transcribed by RNA pol II, these snRNA molecules are not translated into proteins, instead they team up with proteins to form small nuclear ribonucleoproteins (snRNPs) known by the cheerful name of "snurps." They form the spliceosome, which edits messenger RNA after it is transcribed from DNA. Smith, Garrett, and Chengqi Lin, a graduate student in Shilatifard's laboratory, also demonstrated that this function of LEC is highly conserved from Drosophila to mammals.
"The specialization of the SEC and LEC complexes for mRNA and snRNA-containing genes, respectively, suggests the presence of specific classes of elongation factors for each class of genes transcribed by RNA polymerase II, which is of fundamental significance," says Smith.
"The next step is to figure out what other classes of genes use other classes of elongation factors. And what are the differential mechanisms of recruitment to RNA Polymerase II on different classes of genes? Once we get a handle on these distinct classes of genes, we hope to be able to modify different classes of genes by modifying these elongation factors." This would be new perspective regarding basic biology and clinical intervention, Shilatifard believes.
Provided by Stowers Institute for Medical Research