June 20, 2014 report
Solitary acoustic waves observed to propagate at a lipid membrane interface
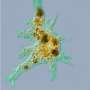
 in lipid interface looks strikingly similar to an AP. Credit: Shamit Shivastava")
(Phys.org) —Defining the essential character of the action potential of neurons has proven to be an elusive task. As typically happens, the biggest advances seem to have been made early on. In this case it was Hodgkin and Huxley working with the giant unmyelinated axons of squids. By squeezing out the "axoplasm" and replacing it with various concoctions these guys could isolate the effects of different ions and the channels through which they flowed. Using their data, they were able to create one of the most successful models known to science.
A couple of concerns remained through all of this, perhaps best highlighted by the ever intriguing and often beguiling Gerald Pollack in his book Cells, Gells, and the Engines of Life. For one thing, membranes seemed to be excitable all by themselves. Pollack notes that patches of bare membrane held within the tip of a patch pipet electrode show tiny "channel" currents even without any protein channels. These heretical mini-spikelets don't seem to be simple leaks at the borders but are more likely transient membrane effects which created tiny pores.
Perhaps even more alarming was the fact that good old axoplasm itself, when stripped of its channel-bearing membrane, still shows some ability to transduce a spike (albeit much attenuated and slowed). Pollack attributes this ability to a propagating phase change in the cytoplasmic gel which unmasks largely immobile negative charges on proteins, previously bound with cations and an ordered hydration shell. This water layer, an interface reaching up to several molecules deep, is also critical to understanding the behavior of the lipid membrane. Some the most novel incites in neuroscience now spring from the thermodynamic characterization of this interface.
In the spirit of physical chemistry a new breed of physical neuroscientists are now combining fairly old, low tech instruments with high tech optics to explore membranes. A recent paper from Shamit Shrivastava and Matthias Schneider in The Journal of the Royal Society publication Interface takes the field a bold step forward with its claims to have caught a glimpse of the elusive creature that has come to be known as the solitary pulse. The mathematical construct known to many as "solitions" are special cases of these more general pulses. This would be the first time that solitary elastic waves have been observed propagating in lipid membranes.
The membranes they studied were monolayers of Dipalmitoylphosphatidylcholine (DCCP), more familiar to us as standard lung surfactant. Most organisms, with exception of a few Archae that have fused two phospholipid tails together, use bilayer membranes which work great for cells. For membrane experiments however, researchers typically use a device known as a Langmuir trough. This workhorse of membrane biophysics (in service since 1917) is basically a small pan with an air-water interface comprised of single amphiphilic monolayer. The membrane can be squeezed from the side as needed, and the lateral surface pressure and membrane tension in the lipid measured with a Wilhelmy plate. This gauge is basically a sensitive electrobalance hooked to vertical plate that is dipped into the trough and wetted. The resultant forces acting on the plate can then be directly quantified.
To initiate longitudinal pulses a razor blade was placed into the trough and actuated horizontally by a piezoelectric element. In some incarnations of the setup a Kelvin or AFM probe can be used to detect surface potential or charge. The refinement that permitted the researchers to remotely sense solitary waves was to use fluorescence resonance energy transfer, or FRET. Here a donor chromophore transfers energy through nonradiative dipole-dipole coupling to an acceptor chromophore when it is within its near field. It can therefore be used as a convenient and fast way to measure the separation between two molecules, and hence their perturbation by a pulse. In ratiometric FRET signal to noise is improved by simultaneously acquiring emission intensity at two wavelengths, here 535 and 605 nm. Critical for these experiments, the longitudinal compression component can be distinguished from the transverse capillary components of a pulse.
Solitary pulses with a threshold of excitation are dependant upon the existence of a nonlinearity in the elasticity of the interface. For both mono- and bilayers this can arise as a peak in their compressibility, or susceptibility (cp,kT, etc.). In a dissipative medium, amplitude decay will eventually result in broadening of the pulse to the point where the nonlinearity can no longer balance the dispersion. At some point the amplitude of the pulse will slip below threshold. For real nerves where spike shape is maintained for long distances, it has been suggested that the pulse is replenished by ion channels along the axons or at their nodes.
Shrivastava says they are now collaborating with Ronald Netz in Berlin to computationally study interstitial sound waves starting from scratch. The properties of the surrounding media play an important role in dissipation and propagation of waves. Of particular interest here is how myelin might aide and abet nerve pulses by virtue of its unique construction or phase transitions of its own. It is also worthwhile to note that cell membranes are not just found at their periphery, cells are filled with them. Transport phenomena, and the trafficking and segregation of proteins through various membrane fluctuations are fascinating new areas of study we might encourage you to read more about on this site.
One prediction of the Hodgkin-Huxely model mentioned above, is that because of channel inactivation, collision of two pulses should result in their annhilation. In many real neurophysiology experiments where so-called "antidromic stimulation" has used to identify which regions of the brain are connected to each other (and how fast the conduction pathways are), this has been found to be the case. However recent studies have found that nerve pulses can and do survive collisions, in agreement with a soliton theory for electromechanical nerve pulse propagation. Although Shrivastava and Schneider found pulse velocities compareable to those of action potentials in unmyelinated axons, they suggest that velocity and pulse shape are not the best criteria to gauge spike propagation. They offer that the variation in velocity as a function of state, the variation of pulse shape as a function of degree of nonlinearity, and the existence of a thermodynamically-defined threshold are the key properties.
There is still a bit of mismatch between this emerging field and traditional biologists who don't typically quantify cells using equations of state. Stepping outward for a moment, we might recall that Schrodinger's wave equation was initially considered by many to be a mathematical fiction. It was eventually concluded that the wave function, meaning a packet of vibrations, depicted probabilities rather than energy. When luminaries like Steven Hawking now speak of a universal wave function—the state function of the entire universe—most physicists will still want more than a psi symbol and a couple more characters. In the same way, biologists want more for the cell.
In starting from the ground up and building membranes from scratch, we now have a way to fully address the spike experimentally. Constructing artificial axon-like geometries from membranes might be the next goal along this path. Adding channels, proteins, subsurface actin or spectrin, and a contiguous internal cytoskeleton will be even more illuminating. If we can then myelinate these structures, as has been already achieved for artificial tubes in a culture setting, we might begin to probe questions famously raised by the late Ichiji Tasaki. A pioneer in measuring all things mechanical and thermal regarding spikes, Ichiji also discovered the insulating function of myelin, saltatory conduction between nodes, and how sound vibrations are transduced into spikes in the auditory system.
Showing that membranes can support solitary elastic signalling is a huge step; demonstrating that they are in fact the physical basis of nerve pulses and communication remains as the challenge.
More information: Evidence for two-dimensional solitary sound waves in a lipid controlled interface and its implications for biological signalling, J. R. Soc. Interface 6 August 2014 vol. 11 no. 97 20140098. rsif.royalsocietypublishing.or … 11/97/20140098.short
Journal information: Journal of the Royal Society Interface
© 2014 Phys.org