New technique sheds light on the mysterious process of cell division

(PhysOrg.com) -- Using a new technique in which models of primitive cells are constructed from the bottom up, scientists have demonstrated that the structure of a cell's membrane and cytoplasm may be as important to cell division as the specialized machinery -- such as enzymes, DNA or RNA -- which are found within living cells. Christine Keating, an associate professor of chemistry at Penn State University, and Meghan Andes-Koback, a graduate student in the Penn State Department of Chemistry, generated simple, non-living model "cells" with which they established that asymmetric division -- the process by which a cell splits to become two distinct daughter cells -- is possible even in the absence of complex cellular components, such as genes. The study, which will be published in the Journal of the American Chemical Society, may provide important clues to how life originated from non-life and how modern cells came to exhibit complex behaviors.
Keating explained that how biological cells split into asymmetrical daughter cells with very different compositions and different "fates" is something of a mystery. Cellular differentiation -- the process by which an unspecialized cell, such as a stem cell, becomes a specialized cell -- requires that different biological components reorganize themselves into each of the resulting daughter cells. For this apparently complex task to be accomplished, some important mechanism must guide both the reorganization of cellular parts and the maintenance of polarity -- the property of a cell to exhibit distinct front and back "sides" with specific placement and distribution of cellular machinery. "Many genes have been implicated in the maintenance of cell polarity and the facilitation of division into nonidentical daughter cells. It's thanks to changes in the expression of these genes that a skin cell becomes a skin cell and a heart cell becomes a heart cell," Keating said. "But our research took a different approach. We asked: In addition to the genetic factors that guide asymmetrical cell division and polarity maintenance, what structural, biophysical factors might be at work, and how might these factors have predated the evolution of the complex genetic systems known to exist in modern cells?"
The team began with the hypothesis that because new daughter cells arise by division of existing mother cells, certain inherited material -- such as the cell membrane -- could serve as a sort of informational "landmark." This landmark could set in motion and guide a cascade of chemical events related to ordered cell division and polarity maintenance. To test this hypothesis, Keating and Andes-Koback built model cells from the bottom up, allowing water, lipids, and polymers to assemble into mimics of the most basic constituents of real, living cells -- such as a membrane and cytoplasm. They then altered the osmotic pressure outside of the "cells" by adding sugar, which forced them to divide in a way that is reminiscent of how living, biological cells split under natural conditions.

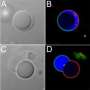
"We observed that even model cells can divide in a structured way, which implies a kind of intrinsic order," Andes-Koback said. She explained that, like a biological cell, the model mother cell was designed to exhibit asymmetry in both its membrane and its cellular interior. The membrane asymmetry was modeled using two distinct lipid domains, while the cellular interior was modeled using two distinct polymers called polyethylene glycol (PEG) and dextran. These polymers form distinct domains, or compartments, on the inside of the model cells, with the dextran-rich compartment containing a higher concentration of a particular protein. The team observed that when the asymmetric mother cell divided, one daughter inherited one lipid domain surrounding the PEG-rich interior, and the other daughter inherited the other membrane domain surrounding the dextran-rich interior, which contained the larger portion of the protein. "Most importantly, we also found that when we varied the relative size of the two lipid domains, one daughter cell got both types of membrane and the other daughter got only one type," Andes-Koback said. "This was possible since the interior aqueous phases controlled the fission plane, and it is important because it provides a way to achieve a patch of distinct membrane to serve as a landmark for polarity in subsequent 'generations.'"
The team members note that the new modeling technique seems to suggests that simple chemical and physical interactions within cells -- such as self-assembly, phase separation, and partitioning -- can result in seemingly complex behaviors – like asymmetric division -- even when no additional cellular machinery is present. "Since there were no nucleic acids nor enzymes present, we clearly didn't have genes governing how our model cells would behave," Keating said. "So our study supports the hypothesis that structural and organizational 'cues' work in concert with genetic signals to achieve and maintain polarity through successive cell-division cycles."
Keating added that a working model of cellular dynamics requires a good understanding, not just of the role of genes, but also of the role of the structural organization of cells. "Once we have a firm grasp of what guides a cell's behavior, we might one day be able to design better disease treatments based on targeting errors in intracellular organization," she said.
Keating also explained that experimentation on non-living model cells that contain no DNA could help point to clues explaining the mysterious process of abiogenesis -- the formation of life from non-living matter, an event that happened at least once during our Earth's history. "Scientists have simulated early-Earth conditions in laboratories and have demonstrated that many amino acids -- the biochemical constituents of proteins -- can form through natural chemical reactions," Keating said. "We hope our research helps to fill in another part of the puzzle: how chemical and spatial organization may have contributed to the success of early life forms."
Provided by Pennsylvania State University