October 4, 2013 report
The machinery of mitosis: Kinetechores, centrioles and chromosome pumps

Credit: University of North Carolina Mitotic Spindle Group
(Phys.org) —At the cellular level, the mitotic spindle apparatus is arguably the most complicated piece of machinery in existence. Its basic function is to isolate and separate the chromosomes during cell division. A group of researchers at the University of North Carolina have been piecing together a model of the spindle and associated proteins which provides a way to visualize in detail exactly what might be going on. The group chose to simulate budding yeast cells because their entire spindle is comprised of only around 40 microtubules (MTs), compared to 100 times that amount in mammalian cells. Over the years the group has contributed to an emerging mechanical picture of the spindle wherein the MTs provide the compression elements, pericentric chromatin the elastic tension elements, and a proteinaceous kinetochore bridges the two polymers together. Their most recent paper, published in Current Biology, provides a new and detailed 3d map of the kinetochore region of the chromosome, and seeks to provides answers to the origins of the seemingly mysterious force that organizes the dividing cell.
Local force generation through the action of motor proteins, like kinesin and dynein, provides an important directional instructor to the spindle. However just as these components are likely not them main source of motility for large organelles like mitochondria, a more holistic view of spindle behavior suggests that a more diffuse, entropic influence constitutes the invisible hand that guides its overall dynamics. One potential source of force generation is provided by the chromosome pumps, if you will, that act like an osmotic contractile gel, expanding and constricting throughout mitosis—perhaps not completely unlike the recently revealed mechanism used in Herpes virus infection. Here the herpes DNA and associated matrix, under extreme pressure, literally blasts itself into the host cell.
The authors note that the entropic elasticity of chromosomal DNA can act to reel the spindle "arms" in to the spindle pole, just as one end of a spring recoils when the other end is pulled to a fxed point. This mechanism of entropic recoil has already been indicated to act in the segregation of replicated DNA in simpler bacteria. One can watch spindle behaviors under the microscope, and see these kinds of mechanisms at work, but they cannot be described very well in paper form by simply detailing lists of binding interactions. So these days, it now seems that in cell biology at least, the model provides the best way to move understanding forward, with parametrically-fed simulations providing the feedstock. Ideally, just like in parametric CAD, if the variables that specify the critical metrics are set up and linked properly, you can change one parameter, and those associated to it seamlessly follow suit without the need to create from scratch.
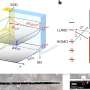
A Matlab/Simulink model was used to generate population measurements of spindle lengths for 16 left and 16 right kinetochore MTs, where each kinetochore has its own associated MT. The entire model contains spindle pole bodies (centrosomes), linked by the kinetochore MTs, with the main barrel structure comprised of coiled DNA tubes. These tubes are stitched together by cohesin rings and condensin linker molecules. In more recent models, the researchers are looking to constrain the unsolved structural details at the surface of the so-called inner kinetochore plate which resides at the kinetochore-MT interface.
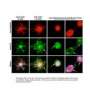
In particular, they are looking to understand the distribution of critical molecules like Cse4 and Ndc80. To this end they created a model based on stochastic growth and shortening of spindles and kinetochore MTs. A cylindrical spindle with a diameter of 250nm was created based on geometry obtained form electron microscopy and tomography. Experimentally, flourophores were placed at the MT plus ends and used to simulate the distribution of Ndc80.
The results of these efforts are not best transmitted in word from. Nowadays collaborations like this are often headed up by artisans that recruit the biology and computer tech experts needed to create a visual simulation. The Harvard Biovisions series, or the illustrative DNA compaction video below are well known examples. The larger public may have to wait a little while to see the fruits of these new labors which seek to make the unseeable biology visible. The mitotic spindle may perhaps be the feature film that many haven't even realized they have been waiting for.
More information: A 3D Map of the Yeast Kinetochore Reveals the Presence of Core and Accessory Centromere-Specific Histone, Current Biology, 26 September 2013. dx.doi.org/10.1016/j.cub.2013.07.083
Abstract
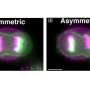
The budding yeast kinetochore is ~68 nm in length with a diameter slightly larger than a 25 nm microtubule. The kinetochores from the 16 chromosomes are organized in a stereotypic cluster encircling central spindle microtubules. Quantitative analysis of the inner kinetochore cluster (Cse4, COMA) reveals structural features not apparent in singly attached kinetochores. The cluster of Cse4-containing kinetochores is physically larger perpendicular to the spindle axis relative to the cluster of Ndc80 molecules. If there was a single Cse4 (molecule or nucleosome) at the kinetochore attached to each microtubule plus end, the cluster of Cse4 would appear geometrically identical to Ndc80. Thus, the structure of the inner kinetochore at the surface of the chromosomes remains unsolved. We have used point fluorescence microscopy and statistical probability maps to deduce the two-dimensional mean position of representative components of the yeast kinetochore relative to the mitotic spindle in metaphase. Comparison of the experimental images to three-dimensional architectures from convolution of mathematical models reveals a pool of Cse4 radially displaced from Cse4 at the kinetochore and kinetochore microtubule plus ends. The pool of displaced Cse4 can be experimentally depleted in mRNA processing pat1Δ or xrn1Δ mutants. The peripheral Cse4 molecules do not template outer kinetochore components. This study suggests an inner kinetochore plate at the centromere-microtubule interface in budding yeast and yields information on the number of Ndc80 molecules at the microtubule attachment site.
Journal information: Current Biology
© 2013 Phys.org