Team solves structure of key mTORC1 activator

A research team led by scientists at the Whitehead Institute has described the structure of a key protein complex that helps activate mTORC1, a master growth regulator that enables cells to quickly respond to changing nutrient levels. This activating complex includes two proteins, folliculin (FLCN), which is disrupted in an inherited tumor syndrome, and FNIP2. As described in the November 5 issue of Cell, the team's findings reveal how FLCN and FNIP2 interact with each other and with other key components of mTORC1 to regulate nutrient and growth signals. These insights will help guide future studies, including the discovery of drugs that interfere with mTORC1 signaling.
"These proteins are a really critical part of the entire mTORC1 system, allowing it to be rapidly turned on in response to the availability of nutrients," said senior author David Sabatini, a Member of Whitehead Institute, a professor of biology at Massachusetts Institute of Technology and investigator with the Howard Hughes Medical Institute (HHMI). "Now, with a detailed understanding of their structures, we can begin to unlock their complex biology."
FLCN and FNIP2 work together to activate mTORC1 but do so indirectly by turning on another set of proteins called the Rag GTPases. This elaborate collaboration has been recognized for several years, but the molecular details—which parts of the various proteins are most important, for example—have remained unclear. In addition, genetic mutations in FLCN are associated with Birt-Hogg-Dube (BHD) syndrome, an inherited condition that includes a predisposition to kidney cancer as well as benign tumors in other parts of the body. How those mutations alter protein function and lead to disease are also unknown.
"We really wanted to understand the underlying mechanistic properties of these proteins to switch on mTORC1 signaling," says first author Kacper Rogala, a postdoctoral fellow in Sabatini's laboratory. "Without knowing the structure, we can make assumptions, but only with a detailed picture can we really understand how they work."
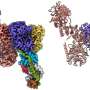
Rogala, together with co-first author Kuang Shen, another Sabatini lab postdoctoral fellow, used cryo-electron microscopy (cryo-EM) to determine the structure of FLCN-FNIP2 in a complex with the Rag GTPases and another key component of the mTORC1 complex (called Ragulator). They worked closely with cryo-EM expert Zhiheng Yu and his colleagues at HHMI.
From these structural studies, Rogala, Shen, and their colleagues made several key discoveries. One unusual finding is that FLCN and FNIP2 bind to each other at two sites—not just one, as is typically observed in complexes comprised of two proteins.
In addition, the two proteins each contain the same two domains: a Longin domain at one end and a DENN domain at the other. The corresponding domains from each protein assemble together, forming what the researchers call "a dimer of dimers."
"This is a very unique protein architecture," says Shen. "In fact, it is quite similar to the structure of another key regulator of mTORC1, called GATOR1." He led a cryo-EM analysis of GATOR1 that was published in Nature in 2018.
Rogala, Shen and their colleagues also identified a key structural feature of FLCN that is responsible for activating the Rag GTPases—a function that relies on enzymatic activity to help switch on mTORC1 signaling. This feature is called an arginine finger because it contains the amino acid arginine at a critical position. Uncovering this structural element will guide efforts to design drugs against it.
Finally, as the team closely examined this arginine finger, they noticed that it wasn't positioned where they expected it to be. In order to activate the Rag GTPases, the arginine finger must be lie very close to the catalytic site—like a key in a keyhole. But in this case, the key is dangling outside of the keyhole.
"This was a real surprise and tells us that we've captured the complex in an intermediate state of some kind," says Rogala. "Now, we can start looking for the other states to examine their structures and begin reconstructing the entire catalytic path."
Now, with a detailed picture of the FLCN and FNIP2 structures, the researchers hope to gain a deeper understanding of the mechanics of mTORC1 signaling—a crucial step toward designing drugs that can potently inhibit these signals. In addition, the team also plans to explore some longstanding biological questions, like how FLCN leads to tumor formation in BHD syndrome.
Proteins that activate mTORC1 are usually considered to be oncogenes, yet FLCN functions as a tumor suppressor—meaning that when its function is lost, tumors can form.
"This has puzzled the field for more than 10 years," says Shen. "There's still no answer, but now, with our structure as a guide, we can begin rationally probing this question—as well as many others—in a much more directed way."
More information: Kuang Shen et al. Cryo-EM Structure of the Human FLCN-FNIP2-Rag-Ragulator Complex, Cell (2019). DOI: 10.1016/j.cell.2019.10.036
Journal information: Cell
Provided by Whitehead Institute for Biomedical Research