What organizes the genome in the nucleus?

Spatial separation of active from inactive fractions of the genome in the cell nucleus is crucial for gene expression control. A new study uncovers leading mechanisms of such separation and turns our picture of the nucleus upside down.
Eukaryotic chromosomes are built of chromatin, a complex of DNA and associated proteins. Depending on transcriptional activity and degree of compaction, two types of chromatin can be distinguished and these two types are spatially separated within the nucleus. The highly condensed fraction is made up of regions of chromatin that contain few genes and is transcriptionally inactive. It is called heterochromatin, and is located in the periphery of the nucleus, close to the nuclear membrane. Euchromatin, on the other hand, is enriched in genes and corresponds to the active fraction of the genome. It occupies the inner regions of the nucleus, is less densely packed, and therefore more accessible to the protein machineries required for gene expression. This general pattern of genome organization is found in virtually all eukaryotic cell types, but the mechanisms establishing the characteristic distribution remain poorly understood. Research carried out by a team led by Irina Solovei at LMU's Biocenter, in cooperation with Job Dekker (University of Massachusetts Medical School) and physicists from the group of Leonid Mirny at MIT (Institute for Medical Engineering and Science) now suggests that the driving force in chromatin segregation is the inactive heterochromatin and that in the 'default' chromatin distribution of euchromatin and heterochromatin are reversed. The new findings appear in the leading journal Nature.
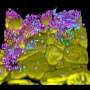
Many mechanisms have been proposed to explain how chromatin is segregated within the nucleus, however none of them were conclusive, largely, because it is difficult to analyze the interactions of the two chromatin types in the context of conventional nuclei with heterochromatin tethered to the nuclear membrane. "For our study, we therefore chose so called inverted cell nuclei," says Solovei. She and her Munich colleagues discovered these nuclei about 10 years ago in the retina of nocturnally active mammals, where they are restricted to the type of photoreceptor cells known as rods. In rods, the tightly condensed heterochromatin is packed in the interior of the nuclei, while the active euchromatin is localized directly under the nuclear membrane—a unique exception to the general rule. It turned out that the heterochromatin core of rod nuclei serves as a microlens condensing light and thus improving optical properties in the nocturnal retinas. Subsequent study from the same group disclosed the mechanism of inversion by revealing that these atypical nuclei lack two protein complexes that normally link the heterochromatin to the inner surface of the nuclear membrane, the nuclear lamina.
 at the top, other retinal neurons with conventional nuclei at the bottom. Credit: I. Solovei")
Using data obtained by a combination of modern microscopy and molecular biology techniques, the researchers have now generated polymer models of the individual chromosomes and of entire nuclei. By simulating the behavior of these polymers under different conditions, they were able to investigate the role of interactions within and between the two chromatin fractions and the nuclear lamina. These studies showed that interactions between heterochromatic regions alone are sufficient for chromatin segregation, whereas interactions within euchromatin are dispensable for this process. "Our results indicate that the inverted nucleus conceptually represents the default nuclear architecture," says Mirny; "while interactions of heterochromatin with the nuclear lamina are essential for building the conventional architecture." "In this respect," says Solovei, "it is intriguing to ask why the majority of eukaryotes have conventional nuclei, and what the functional relevance of heterochromatin positioning at the nuclear periphery might be."
More information: Martin Falk et al. Heterochromatin drives compartmentalization of inverted and conventional nuclei, Nature (2019). DOI: 10.1038/s41586-019-1275-3
Journal information: Nature
Provided by Ludwig Maximilian University of Munich