How zebrafish rebuild the skeleton of amputated fins

Fish, in contrast to humans, have the fascinating ability to fully regenerate amputated organs. The zebrafish (Danio rerio) is a popular ornamental fish. When parts of its tailfin are injured by predators, or are experimentally amputated, the lost tissue is replaced within three weeks. Zebrafish therefore are a favored animal model to study the cellular and molecular principles of organ regeneration.
Zebrafish fins consist of a skin that is stabilized by a skeleton of bony fin rays; similar to an umbrella that is supported by metallic stretchers. Fin rays are formed by bone-producing cells, the osteoblasts. In order to rebuild an amputated fin, a large number of new osteoblasts have to be formed by cell divisions from existing osteoblasts. For this to happen, osteoblasts exposed to the vicinty of the wound have to abandon the production of bone material and revert, or dedifferentiate, to a "rejuvenated" developmental stage. Specialized mature cells, therefore, become osteoblast precursors, which is a requirement to begin several rounds of cell divisions.
Up to now only little was known about how changes in the differentiation status of osteoblasts are brought about, and it was unknown how zebrafish manage to regenerate the exact shape of the lost fin skeleton. Prof. Dr. Gerrit Begemann, at the Developmental Biology unit at the University of Bayreuth (Germany), and Ph.D. student Nicola Blum now report progress on both fronts, published in two articles in the "Advance Online Articles" section of the journal "Development". The new results are likely to inform efforts to reconstitute bone tissue and injured organs in humans.
A cell's dilemma of proliferation versus specialization
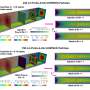
Retinoic acid is required to regulate the addition of bone material in growing fish. During regeneration, mature osteoblasts have to revert to an immature osteoblast precursor, which enables the switch from bone synthesis to cell division. The switch requires retinoic acid levels to drop below a critical concentration. However, upon amputation the tissue beneath the wound initiates a massive bout of retinoic acid synthesis that is required to mobilize cell division in the fin stump. How do mature osteoblasts circumvent this dilemma? The answer was provided by Nicola Blum in the laboratory of Prof. Dr. Gerrit Begemann: Osteoblasts that participate in regeneration transiently produce Cyp26b1, an enzyme that destroys and inactivates retinoic acid. Protected by this process, osteoblasts are able to rewind their developmental clocks, thus turning into precursor cells that contribute to a pool of undifferentiated cells, the blastema. Cells in the blastema pass through a number of cell divisions to provide the building blocks for the regenerated fin.
 of a zebrafish tail fin after amputation. Credit: Developmental Biology, University of Bayreuth.")
However, these cell divisions are supported by high concentrations of retinoic acid, which poses the next predicament: The reversion to become a mature osteoblast is inhibited by high levels of retinoic acid. Nicola Blum found out that connective tissue in those areas of the blastema from which new mature osteoblasts eventually emerge produces the retinoic acid killer Cyp26b1. This lowers the local concentration of retinoic acid, so that osteoblast precursors are again able to mature and produce new fin rays. Other parts of the blastema, which replenish the supply of cells needed for regeneration to occur, continue to produce retinoic acid. "This is an elegant mechanism that ensures a gradient of cells experiencing high and low levels of retinoic acid", Begemann explains, "This allows two processes to run in parallel during regeneration: Proliferation for the production of all cells that replace the lost structure and redifferentiation of osteoblasts where the skeleton re-emerges."
A navigation system that routes cells to regenerating fin rays
How is the exact shape of the fin skeleton regenerated? In order to form new fin rays, newly formed osteoblasts have to align at the correct positions, in this case in extension of existing fin rays in the stump region. The mechanisms ensuring correct osteoblast alignment had remained unknown so far. In a second study, also published in "Development", Nicola Blum broke down the events required for skeletal pattern regeneration.
Osteoblasts are ultimately guided to target regions by a signaling protein called Sonic Hedgehog. This is produced locally in the epidermis, a skin-like layer that covers the fin and the blastema. However, signal production only occurs in locally restricted cells that are free of retinoic acid. Such epidermal cells produce Cyp26a1, an enzyme that is functionally similar to Cyp26b1. By manipulating the levels of retinoic acid metabolism in a way that allows Sonic Hedgehog expression from most regenerating cells, Nicola Blum could show that Sonic Hedgehog acts as a beacon for osteoblast precursor cells. The consequences were dramatic: Instead of aligning with existing fin rays, the cells also invaded the spaces between them, which normally form elastic skin. Eventually bone also formed at inappropriate positions, which in turn sabotaged regeneration and the emergence of the original skeletal pattern.
Lastly, it emerged that osteoblasts themselves exert a piloting function for other cell types, particularly mesenchymal cells and blood vessels that also have to be directed to appropriate destinations during the rebuilding process. If osteoblast precursors are misguided, these cell types follow and exacerbate the inability to reform a functional fin skeleton.
"The re-emergence of the skeletal pattern relies on a navigation system with interacting parts", Begemann summarizes. "Initially, retinoic acid is inactivated where new rays are to form. This allows the local production of a signal that pilots immature osteoblasts to areas where existing fin rays are to be extended. Interestingly, over the course of regeneration other cell types in the blastema are informed by osteoblast precursors to respect the boundaries between emerging fin rays."
More information: Osteoblast de- and redifferentiation is controlled by a dynamic response to retinoic acid during zebrafish fin regeneration. Development 2015, Vol 142 / Issue 17; posted ahead of print August 7, 2015, DOI: 10.1242/dev.120204
Retinoic acid signaling spatially restricts osteoblasts and controls ray-interray organization during zebrafish fin regeneration. Development 2015, Vol 142 / Issue 17; posted ahead of print August 7, 2015, DOI: 10.1242/dev.120212
Journal information: Development
Provided by Bayreuth University