Chitinase as 'burnt-bridge' Brownian monorail efficiently hydrolyzing recalcitrant biomass
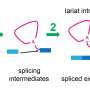
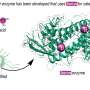
 and chitin binding surface and cleft of SmChiA (right panel). SmChiA moves along with single chain of chitin (shown by red) and cut the chitin (rail) into soluble disaccharides. Credit: NINS/IMS")
Molecular motors convert energy into unidirectional mechanical motion. Most biomolecular motors in cells use adenosine triphosphate (ATP) as a chemical energy source. Recently, however, Serratia marcescens chitinase A (SmChiA) has been rediscovered as a molecular motor working in extracellular environments without using ATP. Similar to a monorail car (Fig. 1), SmChiA has cleft-like polysaccharide binding sites and processively hydrolyzes recalcitrant crystalline chitin, a major biomass source, into a water-soluble disaccharide chitobiose. As a tool of biomass conversion to useful chemicals, SmChiA has been extensively studied.
During the processive catalysis and movement on a crystalline chitin surface, SmChiA binds with single chitin chain in the catalytic cleft and repeats chemical and mechanical steps. In the chemical step, the glycosidic bond is first cleaved and the intermediate structure of substrate is hydrolyzed. The reaction product, chitobiose, is then released, and the next chitobiose unit is peeled from the crystal surface (decrystallization) accompanied with the forward step. Considering the size of the reaction product chitobiose (~1 nm), SmChiA is expected to move with single-nanometer step sizes. Therefore, a single-molecule imaging technique with high precision and speed was required to resolve the single steps coupled with catalysis.
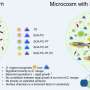
To understand the operation mechanism of fast catalysis and the unidirectional motion of SmChiA, Nakamura and co-workers in the Institute for Molecular Science (IMS) analyzed elementary steps of movement coupled with catalysis using high-precision and high-speed single-molecule imaging probed with gold nanoparticle. They verified fast unidirectional movement (~50 nm s-1) with 1 nm forward and backward steps, consistent with the length of the reaction product chitobiose. Analysis of the kinetic isotope effect revealed that hydrolysis is much faster than decrystallization. The much larger forward-to-backward step ratio is explained by the competition between the catalysis (86 percent) and backward movement (14 percent), indicating that the movement is rectified forward by fast catalysis (Fig. 2). This is the so called "burnt-bridge" mechanism, removing the rail behind, and forcing the molecule to move forward.
. The ratio of catalysis and backward step calculated from time constants were 86 percent and 14 percent, respectively, indicating fast catalysis rectify the Brownian motion along chitin chain (right). Credit: NINS/IMS")
Furthermore, by the collaboration between IMS and Tokyo Institute of Technology, SmChiA was shown to be a "burnt-bridge" Brownian ratchet, verified by X-ray crystallography and molecular dynamics simulation of the intermediate structures during sliding movement. Decrystallization of a single chitin chain is the rate-limiting step of movement achieved by binding free energy at the product binding site, indicated by comparison of free energy differences estimated by the single-molecule analysis with crystalline chitin and theoretical calculation of the binding energy with soluble oligo-saccharide.
The finding demonstrates how SmChiA controls the Brownian motion and extracts fast unidirectional motion for continuous degradation of crystalline chitin without dissociation. The strategy evolved by SmChiA can be applied not only to engineer chitinases and cellulases for more efficient chitin and cellulose degradations, but also to design fast-moving artificial molecular motors such as DNA walkers.
More information: Akihiko Nakamura et al, Processive chitinase is Brownian monorail operated by fast catalysis after peeling rail from crystalline chitin, Nature Communications (2018). DOI: 10.1038/s41467-018-06362-3
Journal information: Nature Communications
Provided by National Institutes of Natural Sciences