Why do cells make invadopodia?
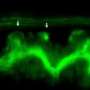
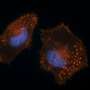
 and basement membrane laminin (GFP) overlayed on DIC. Credit: Lohmer, Clay et al.")
During development and immune system function in animals, specialized cells acquire the ability to invade other tissues. Such invasive behavior is important for neural crest cells that invade skin and provide pigmentation, for embryos that penetrate the uterine wall to establish the fetal-maternal connection, and for immune cells that invade tissues to reach sites of injury and infection. Cell invasive behavior is also acquired by cells in a number of diseases, including asthma, rheumatoid arthritis, and most notably in cancer, where tumor cell invasion results in lethality.
One of the most impenetrable barriers invasive cells must overcome is basement membrane, a dense, sheet-like assembly of proteins and carbohydrates that functions like a wrapping of Kevlar that surrounds and protects tissues. To overcome basement membrane barriers, cells use 'invasive feet' or invadopodia. Invadopodia are like cellular drill bits on the surface of cells that punch through basement membranes. Most cell invasion events are rapid, occur deep in tissues, and are rare. It has thus been difficult to see and study invadopodia in a living animal, even with the most advanced microscopes. Compounding this problem, it has not been possible to perform forward genetic screens to randomly knock-out genes and find those that that control invadopodia formation. As a result, how cells make invadopodia during invasion has been challenging to understand.
How do you study invadopodia?
My lab has been studying how a single cell – the anchor cell – in the nematode worm Caenorhabditis elegans breaks through a basement membrane to connect the uterine and vulval tissues. The C. elegans worm is transparent, and anchor cell invasion is highly stereotyped, allowing invasion to be easily visualized. Furthermore, we have developed different fluorescent proteins that tag molecules that make up the basement membrane and that are found only in the anchor cell, so that we can simultaneously watch basement membrane breaching and molecules in the anchor cell that build invadopodia (see the image above). Finally, we have established screening methods that allow us to rapidly knock down the function of every gene in the worm and see if they contribute to building invadopodia. Using these approaches, in this issue of PLOS Genetics we have performed the first large-scale screen in a living animal to determine what key genes promote invadopodia formation. Our study reports 13 genes as candidates for involvement, and we confirmed that two specifically function to make invadopodia.
 invasion through the basement membrane (BM) into the vulval epithelium in C. elegans. Credit: Lohmer, Clay et al.")
How do the Rho GTPase Cdc42 and the Rab GDP disassociation inhibitor GDI-1 contribute to invadopodia function?
One of the genes isolated was the Rho GTPase Cdc42, a molecular switch that triggers F-actin polymerization. Actin is a cytoskeletal protein whose dynamic polymerization into F-actin (long chains and branched networks of actin monomers) generates the force at cell membranes that drives their protrusion through basement membranes. Using a molecular probe that reports Cdc42 activity we found that Cdc42 is specifically active at the cell membrane of the anchor cell that contacts the basement membrane and seeds the formation of new invadopodia. At any one time, there are approximately 10 invadopodia in the cell that rapidly form and turnover (each invadopodium lives ~45 seconds). Without Cdc42, only about half the number of invadopodia form and these invadopodia are poor invaders and breach the basement membrane in a delayed manner.
We also identified and characterized the Rab GDP disassociation inhibitor GDI-1, which controls the trafficking of intracellular membranes. GDI-1 traffics the specialized invadopodial membrane from within the cell to the site of invadopodia formation at the cell surface. The invadopodial membrane may add membrane to invadopodia so that the invaders can extend through basement membrane, and might deliver proteolytic enzymes that dissolve the basement membrane. In the absence of GDI-1, the invasive cell membrane is mis-targeted to all outer membranes of the cell and fewer invadopodia form.

What's next?
Many questions remain. Loss of neither Cdc42 nor GDI-1 completely blocks invadopodium formation. What other molecule(s) seed invadopodia in the absence of Cdc42? Are the resulting invadopodia molecularly different? – they still breach the basement membrane, albeit in a delayed manner. How is invadopodial membrane delivery to the cell surface coordinated with Cdc-42 F-actin generation to make invadopodia? Does Cdc42 provide a landing spot for the invadopodial membrane? Furthermore, what are the functions of the other 11 genes that promote invadopodia? Characterizing these genes will reveal additional fascinating insights into invadopodia, and will likely lead to new ideas about how to block their formation in human diseases. An important implication from this study is that invadopodia have robust and redundant mechanisms to build them in vivo. Targeting multiple components of invadopodium formation will thus likely be necessary to prevent these structures from forming in human diseases such as cancer.
More information: Lauren L. Lohmer et al. A Sensitized Screen for Genes Promoting Invadopodia Function In Vivo: CDC-42 and Rab GDI-1 Direct Distinct Aspects of Invadopodia Formation, PLOS Genetics (2016). DOI: 10.1371/journal.pgen.1005786
Laura C. Kelley et al. Traversing the basement membrane in vivo: A diversity of strategies, The Journal of Cell Biology (2014). DOI: 10.1083/jcb.201311112
Elliott J Hagedorn et al. Cell invasion through basement membrane: the anchor cell breaches the barrier, Current Opinion in Cell Biology (2011). DOI: 10.1016/j.ceb.2011.05.002
David R Sherwood et al. Anchor Cell Invasion into the Vulval Epithelium in C. elegans, Developmental Cell (2003). DOI: 10.1016/S1534-5807(03)00168-0
Journal information: PLoS Genetics , Developmental Cell
Provided by PLOS Blogs
This story is republished courtesy of PLOS Blogs: blogs.plos.org.