Reverse engineering mysterious 500-million-year-old fossils that confound our tree of life

Paleontologists like us are used to working with fossils that would seem bizarre to many biologists accustomed to living creatures. And as we go farther back in Earth's history, the fossils start to look even weirder. They lack tails, legs, skeletons, eyes…any characteristics that would help us understand where these organisms fit in the tree of life. Under these circumstances, the science of paleontology becomes significantly harder.
Nowhere is this issue more apparent than in the Ediacaran period, which lasted from 635 million to 541 million years ago. A peculiar and entirely soft-bodied suite of fossils from this era are collectively referred to as the Ediacara biota. Despite nearly 70 years of careful study, paleontologists have yet to identify key features among them that would allow us to understand how these organisms are related to modern animals. The forms evident among Ediacaran organisms are, for the most part, truly unique – and we are no closer to understanding their place in evolutionary history.
Rather than looking for characteristics that would allow us to shoehorn some of these organisms into known animal groups, we've taken a different approach. It relies on a technique called computational fluid dynamics that lets us reverse engineer how these organisms lived in their ocean environment.
Mystery fossils
The Ediacaran period marks a pivotal interval in Earth's history; at its start are the last of the so-called "Snowball Earth" events – episodes lasting millions of years when the entire surface of our planet was covered in ice. It segues into the succeeding Cambrian geological period, which saw the first appearance of many of the animal groups we recognize in the present day. This is what's commonly referred to as the Cambrian explosion.
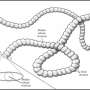
 and Tribrachidium (right). Credit: M. Laflamme, Author provided (No reuse)")
When large, complex fossils were discovered in the Ediacaran, researchers naturally expected that many of them would represent early relatives of the same animal groups that had been recognized in the Cambrian. But these Ediacarans seem completely distinct from modern animals.
For instance, the rangeomorphs were a collection of leaf- and mat-like organisms with a unique fractal architecture, constructed from a series of branching "frond" elements, each a few centimeters long, each of which is itself composed of smaller, identical frond elements.
Another – Tribrachidium – was a small hemispherical organism possessing three raised branches that meet at the top of the organism and which curved toward the margin in a counterclockwise direction.
So how do oddballs like these fit in with what came before and what came after? We just haven't been able to place them on any evolutionary tree.
In order to better understand these organisms, paleontologists have been forced to adopt a different approach. We've abandoned all assumptions about what they might be related to, and instead tried to answer more fundamental questions. For instance, did they move? How did they feed? How did they reproduce? By answering these questions, we can begin to understand their biology and ecology, which in turn may provide hints as to how these organisms are related to other multicellular lifeforms. This is how we've begun to reverse engineer the Ediacara biota.

Modeling fluid dynamics to reverse engineer fossils
One of the most important techniques at our disposal is computational fluid dynamics (CFD), a method for virtually simulating fluid flows around objects using computers.
The rationale for using this approach lies in observing organisms in modern oceans. We know that many (if not all) animals living in shallow marine environments have evolved adaptations that allow them to interact with and manipulate currents, either to reduce drag and prevent them from being swept away (think limpets and barnacles), or to aid in feeding (think crinoids, sea anemones and gorgonian corals). So we can learn a lot about an organism's biology and ecology by studying the way it behaves in moving fluids.
With modern species, researchers can study fluid flows around living animals. But for organisms that have been extinct for over half a billion years – such as the Ediacara biota – virtual simulations using CFD are the only approach.
Here's how we do it. First, we obtain a digital 3-D model of a fossil and place it in a virtual flume tank. Then, we simulate water flowing over and around the digital fossil. Visualizing patterns of flow and recirculation around the organism allows us to test hypotheses about how the organism moved and fed. With something as mysterious and obscure as the Ediacara biota, these insights may bring us closer to understanding what they are.
, with faster flow in red colors, and slow flow in blue colors. Credit: Darroch and Rahman, CC BY-ND")
Our recent work with the enigmatic Ediacaran fossil Parvancorina is an example of this approach. Parvancorina is a simple-looking, shield-shaped organism typically 1-2 centimeters in length, with an anchor-like series of ridges on its top surface. Although it's been interpreted in a variety of ways, most scientists have assumed that it was fixed on the seafloor – what we call sessile. No one has seen any limbs preserved with Parvancorina and it's never been found in association with fossilized tracks or trails.
We decided to test this idea by building 3-D models of the two known Parvancorina species, and then using CFD to see how their unique surface structures affected patterns of fluid flow in different orientations. Our results showed that patterns of water flow around the model were dramatically different depending on the how it was oriented in the current.
Assuming Parvancorina was a suspension feeder, our results demonstrate that it would have been good at capturing the food in the ocean water only when it was oriented in a single specific direction. This is obviously bad news if you're a sessile suspension feeder, like some other members of the Ediacara biota. If you rely on the current to carry water laden with nutrients and food particles to your mouth or feeding apparatus, you want that to happen no matter which way the current is flowing. If you're stuck in one place and the current changes, you've got a problem if you can only gather food when it's coming at you from one direction. Any other plausible style of feeding – for example, scavenging – would also imply these creatures had a mobile lifestyle.
We also used these simulations to calculate drag in different orientations. Although talking about front and back ends in Parvancorina is slightly problematic (because we can't even tell whether it had anything resembling a head and tail), we usually think of the shield end as the front. We showed that the drag experienced by Parvancorina was typically lower when it was placed front-on to current, compared to when it was placed side-on. This is also bad news if you're a sessile organism, because it leaves you open to being ripped from the sediment in strong currents.
The inference from these two observations is clear: Parvancorina was better adapted to life as a mobile, rather than a sessile, organism.

New understanding of Parvancorina lifestyle
This conclusion may sound like a minor footnote in the story of life on Earth. But we believe it has powerful implications for how we view the Ediacara biota as a whole.
First, so little is currently known about Parvancorina that any additional information is crucial. The knowledge that it was mobile will help us work out where this fossil fits in the tree of life.
Second, the inference that Parvancorina was mobile, but nonetheless left no trace of its movement, is important – it means that many other Ediacaran fossils that we've assumed were sessile may actually have been mobile as well. This may require us to reimagine Ediacaran ecosystems as much more dynamic and, by extension, much more complex than we previously thought.
Through using tools like computational fluid dynamics to reverse engineer the Ediacara biota, we're getting closer to understanding what they represent, and how they lived and functioned 15 million years before the Cambrian explosion.
Provided by The Conversation
This article was originally published on The Conversation. Read the original article.![]()