Researchers investigate size, depth, and origin of the living subsurface biosphere

The Sun was once thought to provide energy for all life on Earth - meaning that life could not survive without it. In the 20th century, as astrobiologists began to explore the Earth's most remote and harsh environments, scientists began to question that assumption.
We now know that numerous microorganisms are able to obtain the energy they need for life through chemical reactions that do not involve sunlight. These incredible organisms can be found in many environments – from sediments below the dark ocean floor to microscopic pockets of water inside solid rock.
Many questions about these microbial ecosystems remain. How do microbes get into the deep subsurface in the first place? Are communities capable of growing, or do they just sort of sit there in the rock recycling nutrients and carbon from dead cells? How much of the deep biosphere is actually living, and how much of it is just dead matter trapped in the slow, grinding motion of our planet's geology?
Field studies have revealed that subsurface microorganisms can and do live active lives, even when buried kilometers under the surface. But we're still not entirely sure how large the living subsurface biosphere is, how deep it actually goes, and how it originated.
Previous Studies: A Community Harvest
Organisms in the deep subsurface can be identified by simply digging up samples, sticking them under a microscope, and then seeing what's there. The problem is, even though microbes might be present, it's sometimes hard to tell if they're active – or how they behave in their native environment.
Previously, scientists have tried to define the depth limit for life based on environmental constraints like temperature. In general, the environment gets hotter and hotter as you get closer and closer to the Earth's core. Life simply cannot survive when it gets too hot. However, it's hard to tell just how close to that boundary a living community can get.
"The reality is that in order to live at high temperatures, you are forced to replace your proteins very frequently," said Tullis Onstott, a geoscience professor at Princeton University. "If you do not have enough metabolic energy to support that replacement then you, as an individual cell, cannot live."
High temperature environments can be challenging for life. Cellular components break down at an increased rate. If a cell cannot actively repair the damage, the conditions quickly take a turn toward the uninhabitable. Proteins stop working, causing metabolism to grind to a halt. Cell membranes, cell walls and DNA also begin to deteriorate. So it's not temperature alone that affects habitability, it also comes down to an organism's ability to repair the damage that high temperatures cause.
"You will die at a lower temperature even though under energy and nutrient-rich situations you can live at higher temperatures and to much greater depths," said Onstott. "The most important constraint that this places on deep life is its abundance as a function of depth."
The depth and abundance of living organisms in Earth's subsurface depends on how active they are, and how quickly they can repair and reproduce. This is a question of resources and energy. Previous studies have often focused on the resource part of the question - specifically the resource of organic carbon.
Carbon is a major building block of life on Earth, so microbes need carbon in order to build more microbes. In order to double the number of living cells in the community, the microbes need to harvest enough carbon. Carbon, like most resources in the subsurface, is limited; and a microbe's ability to grab it has to do with its metabolism. One of the principal research foci for deep subsurface life is to utilize biogeochemical models to estimate the in situ metabolic rates and then from these rates estimate the in situ anabolic rate.
"Directly determining the in situ anabolic rate is important for two reasons," said Onstott. "The first is that it tells how quickly cells may be dividing and this is an important parameter when discussing how quickly life is evolving in the subsurface. The second is that it also tells you indirectly about the in situ metabolic rate, which then constrains the biogeochemical rate. The latter is particularly difficult to estimate in hard fractured rock environments."
But here's the problem. Measuring the anabolic rate of microbial communities in situ (e.g. in their natural environment) is extremely difficult. Instead, scientists have traditionally collected microbes from the environment and then grown them in the lab. Once the microbes are happily settled on a petri dish, you can measure their anabolic rate quite easily… but the lab is much different than their natural habitat.
"Any anabolic rate you measure in the lab tends to be a gross overestimate," explains Onstott.
The Energy Angle
In addition to resources, a microbe also requires energy. Energy in the subsurface, where sunlight isn't available, is limited by the amount of minerals or chemicals that can be used in life-sustaining chemical reactions inside the cells. This has given scientists another angle for studying how quickly microbes might reproduce.
In order to estimate the in situ metabolic rates of microbes in the subsurface, scientists have monitored the fluctuation of certain chemicals or minerals involved in the production of energy. Like us, microbes 'eat' things to get energy, and then produce waste. But instead of hamburgers (or veggie burgers if you prefer), a microbe might eat something like molecules of iron oxide. This changes the composition of minerals or chemicals in the subsurface. From these in situ metabolic rates scientists inferred the in situ anabolic rates expressed as doubling times (time required to accrue enough carbon to match that of the biomass).
Scientists first estimated the doubling times of subsurface microbes using this approach in the early 1990s, and they found it to be an incredible 1,400 to 150,000 years in subsurface water at just 200 meters depth! However, in 2005 another study found doubling times - in sediments up to 34 meters below the sea floor to be far less - an average of around 10 months..
A New Method
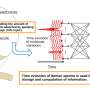
The new study, supported in part by the NASA Astrobiology Institute, took a different approach to the energy angle of subsurface studies. Instead of looking at minerals and chemicals in the environment, they decided to study the damage and repair of molecules inside living cells.

The team developed a method based on this concept that uses the amino acid aspartic acid. Aspartic acid is a chiral molecule, which means it exists in two forms that are "mirror-images" of one another. Living cells produce a lot of 'left-handed' L-aspartic acid, but it also can also exist as 'right-handed,' D-aspartic acid. If life didn't have a preference, there would theoretically be equal amounts of the both L and D forms. But when more L-aspartic acid is found in samples, it's a good sign that you've found an environment with living cells that are making it.
However, environmental pressures are always trying to bring the levels of L- and D- aspartic acid into balance – a process called racemization.
"Of all the amino acids, aspartic acid has the greatest racemization rate," Onstott explains. "Its abundance in prokaryotic proteins is typically 6 to7%, even for subsurface genomes."
This process of racemization is dangerous for cells.
"Prokaryotic proteins (as opposed to peptides in the cell wall) require L-aspartic acid," says Onstott. "If even a tiny fraction of the L-aspartic acid converts to D-aspartic acid, the protein begins to deactivate. As the protein deactivates the substrate that acts as the promoter for this protein will begin to increase, thereby increasing expression of that protein in order to replace the deactivated ones."
Replacing damaged proteins costs a lot of energy and resources. In many Earth environments, this isn't a problem for microorganisms because there are plenty of resources to go around. But for a cell struggling to survive in the deep subsurface, where the local grocery store is always low on supplies or they are difficult to reach, it's a different story.
"Proteins form half of the cellular biomass," said Onstott. "The replacement of deactivated proteins therefore is a significant energy demand on thermophiles in any environment. For certain environments, like hot springs or deep sea vents, the energy flux is sufficient to support this replacement activity to extremely high temperatures. But in the deep terrestrial subsurface, the energy flux is lower."
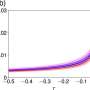
The rate at which racemization of aspartic acid happens can be calculated based on environmental conditions, and it's particularly tied to temperature. You can calculate the amount of damaged proteins a cell will experience over time. You can also calculate the amount of energy they need to replenish damaged proteins. Together, these calculations give you a number for the minimum amount of energy that a microbial biomass needs in order to replace their racemized proteins and survive.
Shedding a Light on the Dark
The new method for estimating microbial survival in the deep subsurface makes significant improvements on previous estimates. Much of this comes down to the fact that scientists are required to make fewer assumptions about the environment.
"Previous methods have an uncertainty of three orders of magnitude," said Onstott.
This uncertainty comes primarily from not knowing what fraction of the total cell population is alive and uncertainties having to do with the biogeochemical rates, the principal metabolic processes, the average cellular mass and the growth yield.
This list of missing data can be filled for other environments where it is easier and cheaper to obtain samples. But sampling the subsurface requires expensive drilling programs or access to deep tunnels where cores can be taken. Even when samples are collected, they're often from a very small section of the local environment (a drill core can be only centimeters in diameter). Because so little is known about the subsurface environment, these assumptions are a major barrier to our understanding of this unique habitat for life.
"Our approach is a direct measure of both the protein doubling time and the active biomass, and does not rely on any of these assumptions," said Onstott. "This approach is particularly powerful for mesophilic (moderate temperature) to hyperthermophilic (extremely hot) environments in the subsurface where it is difficult to constrain the biogeochemical rates."
The Heat of the Deep
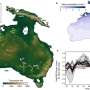
To test their method, the team looked at planktonic microorganisms collected from deep fractures below the Earth's surface in South Africa. Aspartic acid in the samples was heavily tilted toward L-aspartic acid, indicating the presence of living microorganisms.
The researchers found that turnover of amino acid inside cells would take about 89 years at a depth 1 kilometer, where temperatures reached 27°C. At 3 kilometers deep and 57°C, it would take less than 1 to 2 years. This is much faster than previous estimates at this depth based on biogeochemical models.

The numbers also point to the potential for hyperthermophiles living in the subsurface at 85°C.
At 85°C, cells have to replenish their proteins very quickly – about every few days. This is a lot of maintenance for a cell to undertake, and the researchers believe that it might be the cells' ability to perform this protein replacement that limit communities at these depths.
This changes the traditional model of how deep microbes can live on Earth. The resource availability when combined with the requirement for protein replacement limits the number of microorganisms in Earth's deep, hot subsurface.
"This study indicates that the deep biosphere may comprise a smaller portion of Earth's living biomass than originally believed," says Onstott.
Deep Origins and Comparative Planetology
Earth's deep, hot subsurface is a mysterious place. Scientists have only begun to chart the ways in which this biosphere connects to, and interacts with, life at the surface. Some theories suggest that life originated deep inside our planet and made its way above ground over time. Others believe that life originated on the surface and then slowly began to seep into our planet and colonize.
The new study could provide some further insight into these theories. The first point is that a high level of L-amino acids in the deep, hot subsurface (instead of equal L- and D- forms) is likely to be impossible without life.
Even if L-amino acids were produced by natural interactions between rock and water (without life), their formation wouldn't be able to keep up with the rate at which they were being racemized.
This suggests two things. First:
"The development of non-racemic acids and the strong bias for L-amino acids in proteins is something that arose after the origin of life," said Onstott.
Basically, if the origin of life occurred in the deep subsurface, the first proto-cells must have not had a strong preference for L-amino acids because the balance of L- and D- versions would have been relatively equal at that time. Life's taste for L-amino acids would have come later in evolution.
And secondly:
"If non-racemic amino acids are essential for the first proto-cells, then the origin of life could not have occurred in the deep subsurface," said Onstott. "Of course, deep sea vents where you may get formation of non-racemic acids at high temperature and rapid transport into a cold environment remains a viable option."
But even if it didn't begin there, the deep subsurface of Earth may have been a vital refuge for life when conditions at the surface were not as stable as they are today. For instance, the deep subsurface could have sheltered microorganisms from the global devastation of the Earth's surface by large or frequent asteroid impacts. In this way, life could have waited out the disaster in its subsurface bunker, biding its time to thrive another day when the mayhem at the surface cooled down.
The deep subsurface biosphere of Earth likely had a meaningful role in the evolution of our planet, and it also raises questions about the potential for life deep below the surface of other rocky bodies in the Solar System. We now know that liquid water was once present at the surface of Mars, and locations like the Curiosity rover's landing site in Gale Crater were theoretically capable of supporting habitats for life.
If life did gain a foothold on ancient Mars, could it have also gained a foothold in the subsurface of the red planet? And if that were the case, could life still be hiding deep below the martian topsoil, safe from harsh radiation or water sublimation, just biding its time to make an appearance at the surface once again?
The Slow Life
One way that microbes deal with the slow introduction of resources into the subsurface environment is to take the 'slow life' approach. Some microbial communities reproduce very slowly in order to make sure that all of the resources aren't used up at once. br/>
While communities of bacteria on your kitchen counter top might seem to double in size in the blink of an eye, scientists have estimated that subsurface communities might double over hundreds or even thousands of years.
More information: Onstott, T. C., Magnabosco, C., Aubrey, A. D., Burton, A. S., Dworkin, J. P., Elsila, J. E., Grunsfeld, S., Cao, B. H., Hein, J. E., Glavin, D. P., Kieft, T. L., Silver, B. J., Phelps, T. J., van Heerden, E., Opperman, D. J. and Bada, J. L. (2014), "Does aspartic acid racemization constrain the depth limit of the subsurface biosphere?" Geobiology, 12: 1–19. DOI: 10.1111/gbi.12069
Provided by Astrobio.net