Scientists use new simulation methods to observe single events of membrane fusion with molecular resolution
The fusion of membranes is essential for many processes in the human body, for instance, in the communication between nerve cells. A single fusion event occurs on the nanometer scale and takes less than a millisecond. Using computer simulations, scientists at the Max Planck Institute of Colloids and Interfaces have now been able to observe single fusion events with molecular resolution. The simulations are based on new computer algorithms by which one can investigate two large membranes which contain more that ten thousand lipid molecules surrounded by about three million water molecules. These simulations reveal that the fusion process can be controlled by the initial tensions within the membranes. Fusion occurs only at intermediate tensions but, when it occurs, it happens extremely quickly and is completed within 200 nanoseconds.
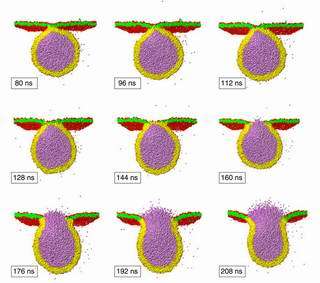
Image: Single fusion event of a vesicle (yellow/orange), that has a diameter of 28 nanometers, with a planar membrane (green/red), that has an area of 50 x 50 square nanometers. The violet particles inside the vesicle represent water molecules; the corresponding particles outside the vesicle are not shown for clarity. The nine snapshots represent the time evolution of the fusion process between 80 and 218 nanoseconds (ns). In this example, the fusion pore has been completed after 160 nanoseconds. Image: Max Planck Institute of Colloids and Interfaces
Membrane fusion is a ubiquitous process of life. A prominent example is provided by the fusion of synaptic vesicles to the outer membranes of nerve cells. This fusion process is responsible for the release of neurotransmitters into the synaptic cleft and, thus, for the communication between these cells. In this way, membrane fusion provides the molecular basis for all our thoughts. Another example is viral infection. Many viruses, such as the influenza or the HIV virus, hide behind a membrane that they have "stolen" from an infected cell. Using this membrane, the viruses can fuse with new cells which become infected as well.
Other examples for membrane fusion include: the fusion of egg and sperm cell during sexual reproduction; the growth of skeletal muscle cells via the fusion of smaller precursor cells (myoblasts); secretion processes such as the release of histamine from mast cells which represents the molecular event underlying many allergic reactions. In addition, we find many more fusion processes when we look at the heavy membrane traffic that is present in all human cells. Indeed, each of these cells contains a huge number of vesicles and other compartments (organelles) that are enclosed in membranes. Many of these vesicles act as transport vehicles that shuttle between the different compartments and deliver their cargo via membrane fusion.
In spite of its ubiquity, the molecular mechanisms underlying the process of membrane fusion are still rather mysterious. There are several reasons for this. First, membrane fusion occurs in the nanoregime and is characterized by length scales between 2 and 20 nanometers. Some experimental tools such as electron microscopy or scanning force microscopy are able to resolve these length scales but only if the membranes are frozen or immobilized and, thus, are no longer able to fuse. Second, a single fusion event represents a relatively fast process. In fact, fusion is so fast that it has so far not been possible to experimentally measure the corresponding fusion time. Third, the fusion of biomembranes involves many different types of molecules, lipids and proteins, which has led to a long and ongoing controversy about the relative importance of these different molecular species for the fusion process.
Using computer simulations, scientists at the Max Planck Institute of Colloids and Interfaces have recently been able to directly observe single fusion events with molecular resolution and to measure the corresponding fusion times. These simulations are based on new algorithms (Dissipative Particle Dynamics - a particle model which allows the simulation of complex fluids at mesoscopic scales) by which one can include a huge number of molecules - in the present case, about ten thousand lipid molecules and about three million water molecules. This makes it possible to study the interaction of a lipid vesicle, which has a diameter of 28 nanometer, with a planar lipid membrane with an area of 50 x 50 square nanometers and a water volume of 50 x 50 x 50 cubic nanometers (see Figure).
When the simulation starts, the vesicle and planar membrane are separated by a thin water layer of 1.5 nanometers. Each membrane consists of a molecular bilayer, i.e., two layers of lipid molecules that are arranged in such a way that the hydrophilic head groups shield the hydrophobic lipid tails from the water. After about 80 nanoseconds (first snapshot in Figure), the membranes have moved into contact by Brownian motion. During the next 60 nanoseconds, the lipid molecules within the contact zone rearrange themselves in such a way that the two membranes undergo hemifusion, i.e., the two bilayers have merged into a single one. This molecular intermingling process seems to represent a new fusion step which has not been reported previously. The resulting hemifused state is rather short-lived since it ruptures relatively fast. This latter step leads to a complete fusion pore that connects the two membranes in a smooth neck--like fashion. The water within the vesicle can then flow through the fusion pore and, in this way, is released from the vesicle compartment.
One important insight provided by these simulations is that the fusion process can be controlled by the initial tensions within the two membranes. For a given membrane, this tension depends on the ratio of the membrane area and the number of assembled lipid molecules. No fusion is observed, within a couple of microseconds, when vesicle and planar membrane are initially relaxed. Instead, the vesicle adheres to and spreads onto the planar membrane. If the vesicle membrane is initially too tense, it ruptures before fusion with the second membrane can occur. Likewise, a large initial tension within the planar membrane leads to premature rupture of this latter membrane. As a consequence, fusion can only occur at intermediate values of the membrane tensions. However, even at those intermediate tensions, only about 55 percent of all fusion attempts lead to successful fusion events. The remaining unsuccessful attempts lead once again to membrane rupture or to stable hemifused states.
Each successful fusion event can be characterized by its fusion time, i.e., by the time from first membrane contact to the appearance of a complete fusion pore. This time represents a random variable since it differs from fusion event to fusion event even if the initial membrane tensions are the same. The statistical distribution of resulting fusion times shows two remarkable features. First, the fusion time distributions for different tensions have significant overlap and are all centered around an average fusion time between 200 and 300 nanoseconds. Second, no fusion event has been observed with a fusion time between 350 nanoseconds and 2 microseconds. This cut-off in the fusion time distribution is related to the stabilization of hemifused membranes at lower tension values.
The fusion of biological membranes is regulated by fusion proteins which are anchored in these membranes. It is generally believed that this regulation is based on conformational changes of these proteins which exert localized tensions and bending moments onto the membranes. In general, different fusion proteins should lead to different force patterns acting on the membranes. Such localized force patterns can be included in the computer models and have been found to induce membrane fusion as well. In fact, using localized force patterns, the fusion process is still characterized by a fusion time in the order of 200 nanoseconds but this process is now more reliable and less random, an obvious advantage for biological fusion processes.
The extension of these simulation studies to multi-component membranes should lead to a deeper understanding of membrane fusion in vivo. It will then be possible to construct biomimetic model systems that are based on the same molecular mechanisms. It seems rather attractive, for example, to use such systems for smart drug delivery. The vision is to enclose the drugs in vesicles that adhere to the unhealthy cells, fuse with these cells, and deliver their cargo only to those cells in a controlled manner.
Original work:
Julian Shillcock and Reinhard Lipowsky
Tension-induced fusion of bilayer membranes and vesicles
Nature Materials (Advanced Online Publication, February 13, 2005)
Reinhard Lipowsky
Biomimetic membrane modelling - Pictures from the twilight zone
Nature Materials 3, 589 - 591 (2004)
Source: Max Planck Institute















