Scientists Reveal a Virus’ Secret Weapon
It takes more than just breaking and entering for a virus to successfully invade a cell. Getting to the cell’s center—where the host cell’s machinery will be co-opted to make more virus—requires navigating obstacles such as membranes and avoiding being recognized and kicked out by the host.
Now scientists from the University of Massachusetts Amherst have uncovered the secret to one virus’s success: a concealed pair of proteins that when unleashed can slice through membranes that are normally impenetrable. The work has implications for fighting a group of viruses that infect the kidneys, respiratory system and brain, and sheds light on how viruses exploit the cellular pathways of their hosts.
The research appears in the Dec. 28 issue of the journal Molecular Cell.
Led by Daniel Hebert, the UMass Amherst team was investigating Simian Virus 40 (SV40), a small, golf-ball shaped virus from the genus known as polyomavirus. Polyomaviruses are nonenveloped viruses—they lack the protective envelope that surrounds some viruses—and SV40 has become the workhorse of scientists trying to understand how nonenveloped DNA viruses work.
Hebert began investigating the SV40 virus to better understand the quality control mechanisms of cells that the virus infects. Comprising only 6 proteins and some DNA, SV40 is what Hebert describes as a “dinky little virus.” Yet, he says, “They know more about the cell than we do—they are surfing down the cellular pathways that we are trying to understand.”
Previous research had detailed part of the virus’ journey to a cell’s interior—SV40 made it into the host cell’s endoplasmic reticulum (ER), a maze-like network of membranes where, among other things, proteins, carbohydrates and fats are put together. But how the virus’ genome got from the ER to the nucleus—a necessary feat because viruses can’t copy their own genetic material and must use the machinery of their host—wasn’t clear.
To investigate, researchers had tried deleting the genes that encoded SV40’s proteins. Of particular interest were the structural proteins known as VP1, VP2 and VP3. Looking at a mature SV40 virus from the outside, one would only see repeated bits of VP1, which make up the exterior of the golf-ball shaped virus. VP2 and VP3 are bound to the interior part of VP1, but on the inside—one could only find them by taking the virus apart. Studies that deleted the genes for these viral proteins had conflicting results—sometimes SV40 was still infectious, in other cases it was not.
So Hebert and his graduate student Robert Daniels decided to couple experiments that knocked out the genes for the viral proteins with experiments that let them follow what exactly happened to virus once inside the host cell’s ER. First his team created mutant SV40s—virus that couldn’t make VP2, virus that couldn’t make VP3 and virus that couldn’t make VP2 or VP3. None of the mutants were capable of infecting cells.
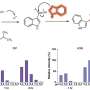
The research team also took a closer look at each of the viral proteins and how they operated, both on their own and as a unit. Generally, proteins are either of the membrane persuasion or they are not—those with the chemical and physical properties that allow them to insert into a membrane aren’t usually found outside of membranes, and proteins with properties that make them adverse to membranes stay out of them. But Hebert and Daniels discovered a strange thing. When VP2 and VP3 were attached to VP1, as they are in the mature virus, none are membrane proteins. But when VP2 and VP3 are on their own, they can slice into membranes, potentially making a pore through which viral DNA could travel to the nucleus.
A protein that can make a hole in a membrane can be a self-defeating weapon for a virus, says Hebert. Slicing membranes before the host’s machinery makes lots more virus means death for the host cell without much viral progeny. But SV40 unleashes V2 and V3 with exquisite timing. The proteins are hidden inside the virus unable to make any holes until they get to the host cell’s ER.
The virus has perfect timing on the construction end as well—once the host starts making more virus, the genes that code for VP1 get turned on first—thus when VP2 and VP3 are made, they can be snatched up by VPI before they can poke themselves through any membranes.
All mature polyomavirus have VP2 and VP3 tucked into their core, suggesting that all polyomavirus use the same secret weapon to gain access to the nucleus, says Hebert. While SV40 doesn’t normally infect humans (it has been found in humans and was likely introduced in the 1950s via a contaminated polio vaccine), its compatriots, the polyomaviruses known as JC virus and BK virus do, and sometimes fatally. These new findings may lead to new strategies for treating or preventing infection by polyomavirus.
Hebert, whose background is in cellular biology, intends to keep studying SV40 to better understand the inner workings of the cells the virus infects. “Viruses provide a valuable window to the cell to follow fundamental cellular processes and we can learn a lot by uncovering how these pathogens exploit their cellular host.”
Source: University of Massachusetts Amherst