June 26, 2020 report
The evolution of the synapse

Among the most easily recognizable features of any nervous system is the synapse. While the question of how synapses evolved has been a longstanding mystery, it can now largely be solved. In a nutshell, it appears that the synapses between neurons evolved directly from the original cell-to-cell contacts, namely, the adherence junctions and other bonds that linked the primitive epithelial sheets of early multicellular organisms.
In other words, the story of how nervous systems originated dates to the very origins of multicellularity itself, or at least close to it. The implications of this assertion are profound. While the exact details are still murky, multicellular organisms evolved through episodes of clonal acquisition and merger. In its simplest incarnation, a dividing cell replicates its DNA and partitions its membrane in the normal way, but the daughter cells remain attached. A crude form of apparent phenotypic differentiation soon follows with the induction of some temporal asynchrony in the maturation processes of the individual cells. Some daughter cells may speed right through to normal adult form while others dawdle about permanently in some new differentiated form.
The other pathway to multicellularity is for independent cells with completely unique DNA to link together somehow. Curiously, this second mechanism persists in sundry extant aggregating organisms like slime molds and choanoflagellates. As a sister group to all metazoans, the choanoflagellates are of special interest here, not just because of their flexible shifts between unicellularity and colonialism, but because of the special cell junctions they developed to control their state and polarity. While aggregation is typically thought of as a response to starvation or stress, there is another critical component that links the behavior of these kinds of organisms more directly to nervous systems: communication.
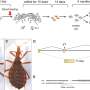
The question of how the communication systems of neurons, namely their neurotransmitter systems, first evolved has now also been largely answered. In a recent sweeping review for Current Biology, Detlev Arendt of Heidelberg, Germany, details the full genesis of the actual hardware of both the pre- and postsynapse. The paper lays down the order in which the canonical small molecule transmitter systems of life sequentially made their first appearance on the stage, and chronicles the novel deployment and proliferation of the associated structural matrix, scaffolding and adhesion molecules that made it all happen.
On the presynaptic side, Detlev subdivides things into three main parts, the active zone vesicle release machinery, the voltage-dependant calcium channels that convert the spike to a calcium transient, and the SNAP/Snare/synaptotagmin complexes that translate the calcium signal into vesicle fusion. In the evolutionary expansion of the synaptotagmins in particular, there seems to have been an important early role in the calcium-regulated transport of glycerolipids between membranes. Once in the form of vesicles, these membrane parcels inevitably also contained hydrophilic proteins, and presumably also soluble peptides esconced or otherwise imported into their interior.
The evolutionary record also shows a suite of vesicular purine nucleotide transporters were present early on in life. With the addition of appropriate receptors to the mix, the makings of crude peptide and nucleotide transmitter systems were already in hand before animals appeared. In choanoflagellates, for example, secretory vesicles bud off the trans-Golgi network to fuse with the apical plasma membrane. A nearly complete calcium-sensitive presynaptic machinery is also active in dense core vesicle secretion in many non-neural secretory cell types.
Aside from the peptide and nucleotide transmitters, the first of the modern amino acid (or amino acid-derived) neurotransmitters appears to be glutamate. Vesicular glutamate transporters in the family SLC17A6-8 (Solute Carrier) are present across all the animals including early-branching sponges and placozoans. The first vesicular inhibitory amino acid transporter (VIAAT-SLC32A1) appears to be that for the uptake of GABA and glycine, as it was present in the cnidarian-bilaterian ancestor. The next to evolve was the vesicular acetylcholine transporter (VAChT-SLC18B3), which has only been found in bilaterians. This gives a rough initial transmitter chronology as follows:
ATP > glutamate > GABA/glycine > acetylcholine
On the postsynaptic side, Detlev notes that the modular structure tends to be much more variable than that of the presynapse. The glutamatergic postsynapse, with its elaborate spine formation, tends to be more complex than the cholinergic postsynapse, although both differ in important ways from the inhibitory postsynapse. For example, receptors directly bind to the scaffolding proteins built around the postsynaptic, density-specific protein Shank in the two excitatory but not inhibitory synapses. The nicotinic acetylcholine receptor family are ligand-gated pentameric ion channels and are distantly related to GABA-A and glycine receptors.
In glutamate postsynapses, the Shank protein connects to the actin skeleton via cortactin, and forms a mesh-like matrix with a scaffolding protein known as Homer. At some point in evolution, it appears that the early glutamatergic postsynapse incorporated an ancient filopodial outgrowth module based on another scaffolding protein known as IRSp53 to establish contact with a presynaptic cell. This union then produced the standard dendritic spine morphology we still see today in many excitatory neurons.
Much of the evidence for this joint venture comes from the IRSp53-containing filopodial-like structures of the so-called microvillar collar found in the apical region of choanoflagellates and sponge choanocytes. In fact, a relic microvillar collar is still critically retained in metazoans in many sensory and secretory epithelial cell types. For example, stable IRSp53-positive microvilli form the business ends of our hair cells, and also that of the spinal-fluid-contacting cells found deep within our cerebral ventricles.
The later-evolving cholinergic postsynapse appears to use the same glutamatergic-style Shank scaffold, unlike the Homer and IRSp53 modules. The inhibitory GABA and glycine postsynapses, on the other hand, do not anchor their ionotropic receptors to each other or the cytoskeleton with Shank, Homer or IRSp53. Instead, they deploy another intriguing molecule known as gephyrin. A curious thing about gephyrin is that it moonlights in another critical, if sometimes enigmatic role: It sits at the apex of a complex molybdenum cofactor synthesis chain, and pops a single Mo ion into the molybdopterin backbone ultimately used in least four human enzymes.
I asked Detlev how this ancient, widely expressed, critical molybdenum synthesis protein might have acquired its synaptic side hussle, and he said, "That's the million dollar question for us, now." Perhaps the clues will soon come from the same kinds of phylogenetic inspection as above. In looking more closely at the many cadherin and integrin proteins that anchor things together at the synapse, it is just now possible to see directly how the rigid apical-to-basal vertical ordering of the specific adherence, occluding and septate junction subtypes found in the most primitive creatures topologically unfolded themselves into linear, polarized, axo-dendritic domains that fix our own nervous systems today.
Perhaps this sentiment is nowhere more poignant than in the exquisitely structured paranode borders of nodes of Ranvier, which serves to facilitate the rapid conduction of nerve impulses in myelinated axon segments. Strictly speaking, tight junctions emerge first in chordates and are located apical to adherens junctions, while the more elusive separate junctions appear on the sides. The funny thing here is that septate junctions have only been unambiguously reported in invertebrates, with the exception of just one place —the vertebrate paranodes.
More information: Detlev Arendt. The Evolutionary Assembly of Neuronal Machinery, Current Biology (2020). DOI: 10.1016/j.cub.2020.04.008
Journal information: Current Biology
2020 Science X Network