How to kill a protein
For decades scientists have been looking closely at how our cells make proteins. But the inverse is equally important: how cells kill them.
While producing healthy, vital proteins is fundamental to life, cells also destroy proteins as a way to switch essential processes on and off. What's more, cells often need to dispose of proteins that malfunction. In fact, protein destruction has emerged as one of the most significant regulatory mechanisms in cells, equal to protein production. There are also many conditions and diseases, such as neurodegeneration, that result when our bodies can't efficiently take out the proteomic trash or destroy a protein in order to flip a critical switch.
We certainly know much more about how proteins are made than we do about how they are unmade, but knowledge of the latter is catching up. Researchers in the lab of Marc Kirschner, chair of the Department of Systems Biology at Harvard Medical School, have recently published twin papers that describe, at the resolution of single molecules, the dynamic process of protein degradation.
The researchers comment that this process bears many similarities to speed dating.
The studies are published in the journal Science.
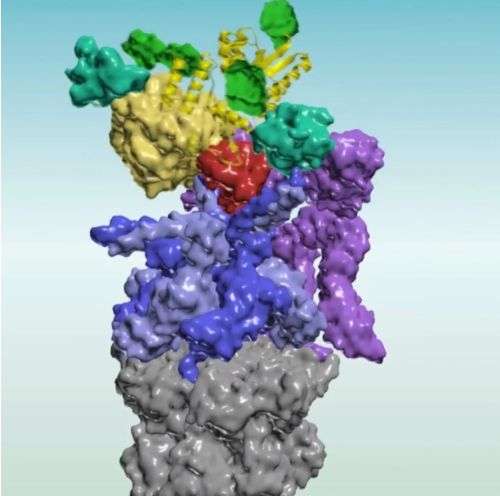
There are many players involved in the act of destroying a protein. A molecular complex called anaphase-promoting complex, or APC, choreographs the intricate events in cell division by sequentially destroying key proteins that block progression of this process. At the right time, APC tags its target with a small protein called ubiquitin. This tagging, a kiss of death, signals a cellular machine called the proteasome.
The proteasome is like a blender stuck on one setting: puree. It is a blunt, and brute, instrument that pulverizes whatever gets inside. Once it smells a ubiquitin tag of a certain type, the protein doesn't stand a chance.
Well, sort of.
Knowledge to date of this process is based on protocols that amass interactions from millions of cells and create static data points that describe a simple, linear process. APC tags a protein with ubiquitin, proteasome shreds the protein. But such data are really no more than a snapshot.
"It turns out that about one-third of all proteins in a cell contain a signal for APC to tag them with ubiquitin, and the proteasome ignores them," said Kirschner, who is also the HMS John Franklin Enders University Professor of Systems Biology. "There's lots of ubiquitin transiently on lots of healthy proteins, in a variety of configurations, for many different reasons and functions. Also, this too is a snapshot since ubiquitin is constantly being added and removed from a protein. So ubiquitin is necessary, but not sufficient. There is far more to the process."
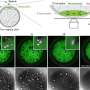
Video above: After a protein attaches to the proteasome, it may tumble between different configurations. The proteasome, in turn, constantly switches its conformation accordingly. These changes create a lock-and-key formation for the protein and proteasome—depending on the ubiquitin chains—which initiates degradation. (Video: Ying Lu)
This led to two basic questions for Kirschner's group: How does APC recognize which proteins to tag, and how does the cell subsequently decide which ones to destroy?
To glean more precise insights into this process, Ying Lu, a postdoctoral fellow in the Kirschner lab, attached individual isolated proteins to a glass slide and exposed them to APC. The proteins, as well as ubiquitin markers, were tagged with fluorescing molecules that enabled Lu to examine individual interactions using a device called a "total internal reflection fluorescence microscope." This allowed Lu to study these interactions one ubiquitin at a time—in real time.
He found that APC doesn't automatically commit itself fully to every protein it encounters. APC, it seems, plays the field.
After all, when someone seeks a mate, they typically don't seek a partner who is merely suitable—they want the perfect match. Likewise, APC doesn't commit to the first protein it dates. Rather, it samples the protein population. It will tag a few proteins here and there, testing for affinity, then move on and try out a few others. Often, it takes only one or two encounters for APC to realize it's not interested. With other proteins, though, it might return and deposit a few more tags, taking things a little further. The more affinity it senses, the more visits it pays.
And the ubiquitin tags accumulate. Once APC has sufficiently narrowed its selection, the tags form a chain that signals the proteasome.
"This approach allows APC to sort through a large number of potential partners rapidly to get to the ones it really wants," said Kirschner.
But this led Kirschner and Lu to wonder what, exactly, the proteasome is looking for. After all, there is a vast range of complex shapes these chains can form, and the researchers suspected that this structural variability was essential to the process.
Since the sheer number of potential structures a ubiquitin chain can form is large, Lu decided to synthesize a variety of shapes in order to get at the basic rules. As he assembled the various data points about structure, he discovered that this was a two-step process.
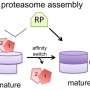
Because the degradation machinery in the proteasome never switches off, it can be a dangerous machine. To protect healthy molecules, the proteasome is sealed with a protective cap, much like the cover of a blender.
Lu found that the first step in destroying a protein was simply to get the molecule to bind to the proteasome's cover. This appeared to be fairly straightforward, based on ubiquitin copy numbers alone. When a protein acquired a particular threshold of ubiquitin, it adhered to the proteasome.
But the second step, to actually get inside of the machine, was far more complex. Lu found that this was by no means a simple matter of opening and closing the lid. Rather, he discovered a dynamic process in which the proteasome's outer surface changes shape, shifts into new positions and actually wiggles itself into a new configuration in order to pull the protein inside.
In other words, the proteasome's protective layer seems to exist in a permanent state of change.
"The dynamics are the whole story," said Kirschner. "Specificity lies in the dynamics."
"We have come a long way in biology looking at static structures, and we've learned a lot," continued Kirschner. "But biology is half knowing what the static picture is and half knowing the dynamics. Researchers have studied the process of protein degradation at a static level for a long time, yet none of this dynamic information on the molecular level was knowable. Now we are developing the tools to look at these processes dynamically. This is a new era."
More information: "Substrate degradation by the proteasome: A single-molecule kinetic analysis." Science 10 April 2015: Vol. 348 no. 6231 DOI: 10.1126/science.1250834
"Specificity of the anaphase-promoting complex: A single-molecule study." Science 10 April 2015: Vol. 348 no. 6231 DOI: 10.1126/science.1248737
Journal information: Science
Provided by Harvard Medical School