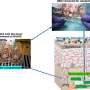
Team applies new theory to learn how and why cells differentiate

How does a stem cell decide what path to take? In a way, it's up to the wisdom of the crowd.
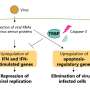
The DNA in a pluripotent stem cell is bombarded with waves of proteins whose ebb and flow nudge the cell toward becoming blood, bone, skin or organs. A new theory by scientists at Rice University shows the cell's journey is neither a simple step-by-step process nor all random.
Theoretical biologist Peter Wolynes and postdoctoral fellow Bin Zhang set out to create a mathematical tool to analyze large, realistic gene networks. As a bonus, their open-access study to be published this week by the Proceedings of the National Academy of Sciences helped them understand that the process by which stem cells differentiate is a many-body problem.
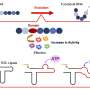
"Many-body" refers to physical systems that involve interactions between large numbers of particles. Scientists assume these many bodies conspire to have a function in every system, but the "problem" is figuring out just what that function is. In the new work, these bodies consist not only of the thousands of proteins expressed by embryonic stem cells but also DNA binding sites that lead to feedback loops and other "attractors" that prompt the cell to move from one steady state to the next until it reaches a final configuration.
To test their tool, the researchers looked at the roles of eight key proteins and how they rise and fall in number, bind and unbind to DNA and degrade during stem cell differentiation. Though the interactions may not always follow a precise path, their general pattern inevitably leads to the desired result for the same reason a strand of amino acids will inevitably fold into the proper protein: because the landscape dictates that it be so.
Wolynes called the new work a "stylized," simplified model meant to give a general but accurate overview of how cell networks function. It's based on a theory he formed in 2003 with Masaki Sasai of Nagoya University but now takes into account the fact that not one but many genes can be responsible for even a single decision in a cellular process.
"This is what Bin figured out, that one could generalize our 2003 model to be much more realistic about how several different proteins bind to DNA in order to turn it on or off," Wolynes said.
A rigorous theoretical approach to determine the transition pathways and rates between steady states was also important, Zhang said. "This is crucial for understanding the mechanism of how stem cell differentiation occurs," he said.
Wolynes said that because the stem cell is stochastic—that is, its fate is not pre-determined—"we had to ask why a gene doesn't constantly flip randomly from one state to another state. This paper for the first time describes how we can, for a pretty complicated circuit, figure out there are only certain periods during which the flipping can occur, following a well-defined transition pathway."
In previous models of gene networks, "Instead of focusing on proteins actually binding to DNA, they just say, 'Well, there's a certain high level of this protein or low level of that protein,'" Wolynes said. "At first, that sounds easier to study because you can measure how much protein you've got. But you don't always know if it is bound. It has become increasingly clear that the rate of protein binding to DNA plays an important role in gene expression, particularly in eukaryotic systems."
The notion that many-body effects even existed in cells began in 1942 when British scientist C.H. Waddington established the idea of an epigenetic landscape for stem cells as a way to describe why pluripotent cells in embryos are destined to turn into bone, muscle and all the other parts of the body – but don't turn back. Waddington compared the cells' paths to marbles rolling to the bottom of a valley.
That concept rang true to Wolynes. His energy landscape theory has become key to understanding protein folding, although that theory sees the landscape as a funnel rather than a valley. "Waddington said that as a cell develops into an embryo and beyond, it becomes many different kinds of cells," Wolynes said. "Those cells might branch off and differentiate further, but they don't typically go back to the original state and start over.
"His analogy – the idea of falling down through a valley – kicked around for a long time, but it was hard to make it mathematically precise. In his time, they didn't know about DNA," Wolynes said.
In both energy and epigenetic landscapes, Wolynes said, the steady state at the bottom is an attractor. "It means wherever you start from, you end up attracted to that same place," he said. "In genetic networks, things like steadily oscillating patterns can also be considered attractors."
Once biologists began to understand genetic switches in DNA, the whole picture became more complicated, he said. "The landscape now has to incorporate the active parts of DNA that are trying to decide whether to turn this gene on or that gene off. In the '50s, we learned how genes made decisions on the basis of their production of proteins. These proteins then act back on the same genes in a kind of feedback loop."
The loops allow genes to remain active for far longer than it would take a protein simply to bind or unbind to a section of DNA. In the researchers' equations, the loops become attractors that help regulate transformation of the cell and can be mapped onto the many-body landscape.
Analyzing the coupled dynamics of all these chemical reactions in a cell could be done by brute force, he said, but the computational cost would be enormous. So the Rice team decided to take a big-picture approach based on Wolynes' earlier work. It so happened that the resulting theoretical models of embryonic stem cells matched nicely with what experimentalists had seen in their studies.
For example, the models explained the fluctuations experimentalists had observed in the expression of a master regulator, a protein called nanog, and its important role in maintaining a cell's pluripotency. Stem cells move from one steady state to the next on their journeys; in their calculations, they found a much higher level of nanog gene expression in what they called SC1, the basic stem cell, than in SC2, a stem cell that had moved to the second steady state. This matched what experiments had measured, the researchers said.
"This is still just a beginning," Wolynes said. "We're looking at embryonic stem cells now, but someday we want to treat the complete developmental program of organisms with hundreds of genes. We can see how these mathematics can scale up to that regime."
More information: tem cell differentiation as a many-body problem, PNAS, 2014. www.pnas.org/cgi/doi/10.1073/pnas.1408561111
Journal information: Proceedings of the National Academy of Sciences
Provided by Rice University