The turn of the corkscrew: Structural analysis uncovers mechanisms of gene expression
The diverse functions of living cells are all based on the information encoded in the structure of the hereditary material DNA. Gene expression must therefore be tightly controlled, and this task is accomplished by the binding of regulatory proteins to, and their removal from, specific DNA sequences. One class of large molecular machines known as Swi2/Snf2 remodelers plays a central role in modulating these processes. However, until now, it was not clear how Swi2/Snf2 remodelers actually work.
A team led by Professor Karl-Peter Hopfner at the Gene Center at Ludwig-Maximilians-Universität (LMU) in Munich has clarified the structure and function of the remodeler Mot1 (Modifier of Transcription 1), which binds directly to DNA. It turns out that Mot1 acts like a molecular corkscrew that migrates along the DNA, following its helical contour. During its progress, Mot1 displaces a crucial transcription factor called TBP (for "TATA Box Binding Protein") from the DNA. Removal of TBP from a TATA box represses transcription of the adjacent gene, and the protein encoded by that gene is no longer synthesized. At the same time, TBP is stabilized and its binding specificity is changed, which facilitates the expression of genes that lack TATA boxes and code for other proteins. (Nature 6 July 2011)
The DNA in the cells of higher organisms is tightly wrapped around protein complexes called nucleosomes. This type of structural organization not only makes it possible to package the long DNA molecules in a highly compact form, it also provides the basis for the controlled expression of genetic information. Densely packed sections of the molecule are effectively in a repressed state, and genes located in these DNA segments cannot be transcribed. Activation of repressed genes depends on the intervention of complex molecular machines, so-called Swi2/Snf2 remodelers, which reorganize condensed stretches of DNA so as to make them accessible for transcription. The precise mode of action of remodelers has so far been unclear, mainly because most of them are made up of several components and the active complexes are difficult to study. This is why Hopfner chose to study Mot1, which is a comparatively simple representative of the family that functions as an Swi2/Snf2 remodeler on its own, and can serve as a guide to understanding the more complicated members of the class. Mot1 is known to participate in the control of gene expression, but how exactly it does so is not well understood.
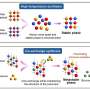
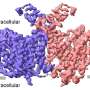
The first stage of the process that leads to the synthesis of a given protein is the transcription of the specific segment of DNA that codes for it into molecules of messenger RNA. This initial step requires the action of so-called transcription factors. One of the most important of these is TBP, which binds preferentially to DNA sequences called TATA boxes that are located near the beginnings of many genes. Binding of TBP introduces a kink into the DNA, and this landmark serves as a platform for the binding of further proteins, ultimately leading to the assembly of the complex necessary for the initiation of transcription. Mot1 regulates transcription by actively removing TBP from the DNA, using ATP as a source of energy. "How Mot1 dissociates the TBP-DNA complex was completely unclear up to now," says Hopfner. With the aid of so-called hybrid methods - in which data obtained from high-resolution X-ray diffraction analysis of the crystallized protein complex with images of the same molecular complex taken with the electron microscope were combined - Hopfner's team was able to define the three-dimensional structure of the Mot1-TBP complex for the first time. This revealed how Mot1 recognizes the surface of the DNA-bound TBP. "Once Mot1 has recognized TBP, it binds to the adjacent DNA and begins to migrate along the DNA strand, using the energy released by the hydrolysis of ATP to power its movement. This helical movement, which is reminiscent of the insertion of a corkscrew, causes TBP to detach from the DNA," explains Dr Petra Wollmann, who is first author on the new study. The researchers were surprised to find that Mot1 contains a strikingly extended loop. After TBP has dissociated from the DNA, this loop masks TBP's DNA binding site and prevents the protein from reoccupying it.
Previous studies had reported what appeared to be paradoxical observations, which indicated that Mot1 inhibits transcription of TATA box-containing genes while facilitating the expression of genes that lack canonical TATA boxes. "Our results suggest that Mot1 also stabilizes the DNA-free conformation of TBP, increasing the probability that it can reach, bind to and activate genes that lack TATA boxes," explains Hopfner. In other words, Mot1 is also a redistribution factor, which enables TBP to bind to different sequences and thus controls its association with other cellular components. This combination of detachment and redistribution functions may be a common feature of remodeling complexes, and would help to explain how they mediate the large-scale redistribution of DNA-binding regulatory proteins.
Provided by Ludwig-Maximilians-Universitat Munchen