Protein clamps tight to telomeres to help prevent aging... and support cancer
The number of times our cells can divide is dictated by telomeres, stretches of DNA at the tips of our chromosomes. Understanding how telomeres keep our chromosomes - and by extension, our genomes - intact is an area of intense scientific focus in the fields of both aging and cancer. Now, scientists at The Wistar Institute have published the first detailed report on the structure and function of a crucial domain in the protein known as Cdc13, which sustains telomeres by clamping to DNA and recruiting replicating enzymes to the area.
While the nature of this portion of Cdc13 had previously eluded scientists, the Wistar researchers found that two copies of the protein bind together to form what is called a "dimer," and how that dimer physically interacts with DNA, regulating how enzymes called telomerases access and lengthen the telomeres. The study was performed using the yeast gene, however, this essential life process has changed little through evolution, and evidence suggests that the human equivalent of this protein may make a good target for future anticancer drugs. They present their findings in the journal Molecular and Cellular Biology, available online now, ahead of print.
"Cdc13 has a crucial support role in maintaining and lengthening telomeres, which are reduced in length through every round of DNA replication," said Emmanuel Skordalakes, Ph.D., assistant professor in Wistar's Gene Expression and Regulation Program and senior author of the study. "We know that disabling this protein in humans will most likely lead to senescence, which is of particular interest in cancer, because telomere lengthening is one of the ways cancer cells obtain their immortality."
In the present study, Skordalakes and his colleagues detail how Cdc13 serves a dual function in telomere replication. First, it keeps the cells' natural DNA repair mechanisms from mistaking the telomere for a broken stretch of DNA, which could cause genetic mayhem if such repair proteins fuse the ends of two chromosomes together, for example. Secondly, Cdc13 recruits telomerase and related proteins to place in order to lengthen the telomeres.
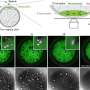
When the researchers introduced mutations into Cdc13 that prevented the protein from forming a dimer, it caused the telomeres to shorten, which would hasten the demise of the yeast cells. When they created mutations that prevented Cdc13 dimers from binding to DNA, it had the effect of excessively lengthening telomeres, an act the researchers attribute to the notion that Cdc13 helps regulate the ability of DNA-replication enzymes to access telomeres.
"The complex role of Cdc13 underscores the unique nature of telomeres and the fine balance between normal cell division and cancer," said Skordalakes.
Telomeres are important to cell division because they serve as sort of a timing mechanism that can, in effect, limit the number of times a normal cell can divide. As each cell divides, it must first replicate - or copy - the DNA of its chromosomes in exacting detail.
However, the proteins in cells that make this replication possible physically cannot copy the last few base units of DNA at the tips of the chromosomes, which effectively shortens the telomere each time a chromosome is copied. Without telomeres to serve as a buffer, a chromosome could conceivably lose a functioning gene as it is copied. This natural "lifespan" of cells was first identified in the 1960s as the Hayflick Limit, named after its discoverer, Leonard Hayflick, Ph.D., then a Wistar scientist.
In 2008, the Skordalakes laboratory was the first to determine the 3-D structure of the catalytic subunit of the enzyme telomerase, which functions to tack on the short stretches of DNA at the telomeres that the cell's main DNA-replicating enzymes miss. The act of preserving telomeres through telomerase is a hallmark of only certain cells, particularly those in developing embryos. In adults, telomerase is active in stem cells, certain immune system cells and, most notably, cancer cells.
According to Skordalakes, the discovery of the dimeric nature of Cdc13 sheds light into the core function of this protein, the recruitment of telomerase (which is also a dimer) to the telomeres. Within the Cdc13 dimer are multiple sites that can bind to DNA with varying degrees of affinity. This allows Cdc13 to straddle the DNA so that one section grips tightly to DNA, while another section - with a more relaxed grip - can bind nearer the tail end of the DNA strand and where telomerase binds. This feature of Cdc13 also assists in recruiting telomerase, summoning the enzyme into place above the telomere.
"You can think of Cdc13 as if it were you hanging on to the edge of a cliff, with one grip stronger than the other," Skordalakes said. "You're going to keep that strong hand on the cliff's edge while your weaker hand reaches into your pocket for your phone."
When Cdc13 interacts with telomerase, Skordalakes says, its weaker hand lets go of DNA, allowing the telomerase to access the telomere while the "strong hand" keeps the telomerase-Cdc13 complex firmly attached to the chromosome end. "It effectively serves as both a protective placeholder and a means of guiding telomerase activity," Skordalakes said.
The Skordalakes laboratory continues to explore the complex biology of telomeres, as well as the numerous other proteins necessary for telomere lengthening to occur. Meanwhile, they are investigating the potential of small molecule inhibitors to serve as viable therapeutics against cancer by blocking telomerase and their related proteins.
Provided by The Wistar Institute